Использование снежного покрова тетеревиными птицами.
В каждой из этих групп морфологические адаптации играют большую, но далеко не единственную роль. Наряду с ними в успешном решении задач, связанных с питанием и уменьшением тепловых затрат, особенно важным становится поведение птицы. И здесь прежде всего особого внимания требует использование снежного покрова для сооружения в нем специальных термических убежищ — подснежных камер. Тетеревиные птицы единственные в классе птиц, которые приобрели способность создавать такие убежища и регулярно пользуются ими в течение всей зимы. Подснежные камеры сооружаются и используются всеми за одним исключением (Bonasa sewerzowi) видами семейства, хотя регулярность их использования зависит прежде всего от степени суровости зимы и характера снежного покрова.
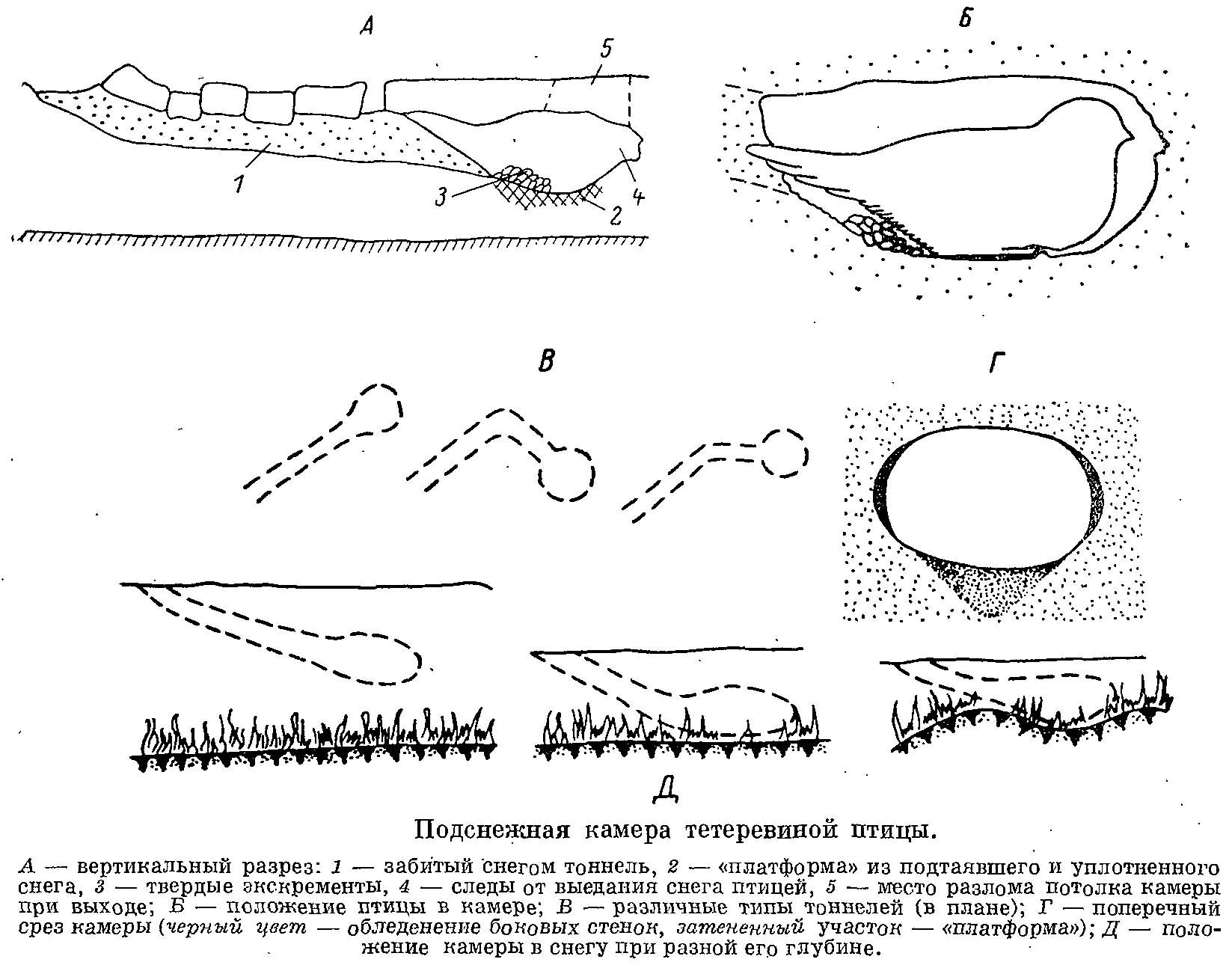
Рис. 1. Подснежная камера тетеревиной птицы. А — вертикальный разрез: 1 — забитый снегом тоннель, 2 — «платформа» из подтаявшего и уплотненного снега, 3 — твердые экскременты, 4 — следы от выедания снега птицей, 5 — место разлома потолка камеры при выходе; Б — положение птицы в камере; В — различные типы тоннелей (в плане); Г — поперечный срез камеры (черный цвет — обледенение боковых стенок, затененный участок — «платформа»); Д — положение камеры в снегу при разной его глубине.
Подснежная камера — это полость в снегу, вырываемая самой птицей (рис. 1). Размеры камеры несколько превышают размеры самой птицы, что дает ей возможность распушить оперение для повышения его термоизолирующих качеств. Толщина снежного потолка может быть различной в зависимости от толщины и структуры снежного покрова, но она всегда выбирается с таким расчетом, чтобы птица могла, встав на пол камеры, высунуть голову наружу. Необходимость в этом существует постоянно, как для регуляции температуры в камере, так и при ее оставлении для предварительного обзора местности. Толщина потолка может резко возрасти, если после зарывания птицы снегопад будет продолжаться всю ночь. Это может побудить птицу покинуть камеру раньше времени. Пол камеры по сравнению со стенками значительно уплотнен и имеет легкий наклон вперед, отчего сидящая в камере птица располагается слегка грудью вниз. Как повышенная теплоотдача через пальцы птиц, так и выделяемые птицей твердые экскременты способствуют некоторому подтаиванию пола, уплотнению и смерзанию находящегося под ним снега. Стенки камеры почти не обледеневают, и пористость окружающего камеру снега практически не нарушается, т. е. его теплоизолирующие качества не меняются. В передней части камеры почти всегда видны углубления и борозды в снегу, оставляемые клювом при поедании птицей снега.

Рис. 2. Время пребывания тетеревиной птицы в подснежной камере (ч в сутки) в зависимости от атмосферной температуры. По оси ординат — часы суток; по оси абсцисс — атмосферная температура, °С.
Строительство подснежной камеры для сна.

Таблица 1. Энергетический выигрыш, получаемый тетеревиными птицами в результате пребывания в термическом убежище при атмосферной температуре —20°С.
Прежде чем соорудить подснежную камеру, птица, сев на снег, сначала прокладывает в нем ход — тоннель, длина которого у разных видов, а также у одного и того же вида может варьировать от 0.2 до 4 м. Наиболее длинные ходы делаются рябчиком B. bonasia, но далеко не всегда. Подснежный ход бывает обычно прямым, но в большинстве случаев перед входом в камеру он делает поворот на 80—90°. Проделывая такой ход, птица, видимо для контроля глубины, один-два раза, а то и несколько раз высовывает голову на поверхность снега. Если ход проделывается в рыхлом свежевыпавшем снегу, то птица просто проминает в нем ход своим телом, в более же твердом слежавшемся снегу она активно работает лапами, клювом, а в начальных стадиях погружения и крыльями. В отношении техники зарывания, птиц в снег долго преобладали совершенно неверные представления, согласно которым птицы просто падали в снег, погружаясь в него под силой тяжести, и дальше уже начинали зарываться активно. Детальные наблюдения на этот счет, выполненные в последнее время на разных видах (Prawdzik, 1963; Pauli, 1974; Потапов, 1974; Андреев, 1975а), показали, что такого пассивного «падания» в снег, да еще со сложенными крыльями, никогда не происходит. На открытую поверхность поляны или болота птица действительно садится с хода (если снег достаточно мягкий) и по инерции делает в таких случаях небольшую ямку с холмиком снега перед ней, взрыхленным при посадке («ударная волна»). Но после этого само погружение в снег происходит уже активно. В большинстве же случаев прежде чем зарыться в снег птица садится на его поверхность и проходит некоторое расстояние пешком. Это особенно характерно в тех случаях, когда поверхность снега достаточно затвердевает и приходится искать места, где снег более рыхлый. Снег слеживается и уплотняется под действием ветра особенно сильно на достаточно открытых пространствах, но среди лесной и кустарниковой растительности сохраняет свою рыхлость значительно дольше. Заваленные снегом кустарники, кроме того, своими ветвями препятствуют его уплотнению. И, наконец, в некоторых случаях птицы могут использовать следы лосей, пробивающих твердые слои снега (мои наблюдения в Ленинградской обл.).

Таблица 2. Биоэнергетика тетеревиных птиц при атмосферной температуре —20 °С.
Само зарывание происходит путем активной работы лап, а в дальнейшем, при продвижении под снегом — и клюва. В процессе рытья птица отбрасывает снег лапами назад, и этот снег забивает просвет тоннеля позади птицы, но не полностью — снежная кровля над ним несколько проседает и ход тоннеля под снегом хорошо заметен на его поверхности в виде небольшого желобка. В свежевыпавшем рыхлом снегу камера в значительной степени обминается, а не отрывается. Если снег достаточно рыхлый, то птица на первых стадиях закапывания может действовать и крыльями, забрасывая снег на спину. Это особенно характерно при зарывании птиц с целью избежать опасности (Prawdzik, 1963). Само зарывание в снег происходит исключительно быстро: тундряная куропатка тратит на приготовление подснежной камеры 15 с (Андреев, 1975а), полевой тетерев — около 1 мин (Pauli, 1974).
Главный выигрыш, получаемый тетеревиной птицей в подснежной камере, — резкое снижение энергетических расходов на терморегуляцию. Хорошие теплоизоляционные качества снега позволяют птицам создавать и поддерживать в подснежной камере температуру, в общем-то оптимальную в данных условиях. Как показали экспериментальные исследования (Волков, 1968а; Андреев, 1977б), температура в подснежной камере практически не зависит от наружной температуры воздуха, держась в пределах —4-—5 °С даже в морозы —40-—50 °С. Оптимальность такой температуры обусловливается тем, что при более низких температурах возрастают расходы на терморегуляцию, а при более высоких создается опасность перехода температуры через нулевой порог, таяния снега и намокания оперения птицы. Отрыв камеру, птица обычно очень быстро, в течение получаса, разогревает ее до указанных температур, чему помогает небольшой объем камеры. Постоянное выделение тепла отдыхающей птицей, в том числе и тепло от твердых экскрементов, не только поддерживает эту температуру, но и создает опасность перегрева, и при повышении температуры выше опасного предела птица проделывает в потолке вентиляционное отверстие (Андреев, 1977б). Все же в ряде случаев температура здесь поднимается выше 0 °С, о чем свидетельствует некоторое обледенение стенок камеры. Такое повышение температуры, видимо, не представляет опасности для птицы в условиях сухого морозного снега, адсорбирующего образующуюся влагу. В последнее время это было доказано экспериментально. В подснежных камерах тетеревов и глухарей температура колебалась ночью от —2 до +3 °С (Marjakangas et al., 1983). Эти же авторы обнаружили слабую гипотермию у глухаря, снижавшего ночью температуру тела в камере с 41.1 до 39.3 °С. Полевой тетерев снижал температуру тела в тех же условиях только на 0.5 °С. Примечательно, что у глухарей, ночевавших на поверхности снега, температура тела снижалась ночью только на 0.5 °С, как и у тетерева в подснежной камере.

Глухарь в снегу.
Физиологические изменения птиц в зимний период.
Высокая степень приспособленности морфологии и физиологии тетеревиных птиц к длительному пребыванию в подснежной камере проявляется буквально во всем. Роговая бахрома на пальцах резко увеличивает эффективность их роющей деятельности в рыхлом снегу. Густое и рыхлое оперение плюсны и нижней части брюха создает теплую, с хорошими теплоизоляционными качествами подстилку, на которой покоится тело птицы при нахождении в подснежной камере. Рассученность, волосовидность оперения в этих местах, имеющего к тому же жировую смазку (обнаруживаемую на ощупь), препятствует даже частичному намоканию его при некотором подтаивании снежного пола камеры от тепла пальцев лап и выделяемых экскрементов. Высокое теплоизоляционное качество всего оперения, необходимое при морозе, крайне выгодно и здесь, в подснежной камере, препятствуя ее излишнему перегреву. В узком замкнутом пространстве подснежной камеры опасна и повышенная влажность — и действительно она здесь не образуется. Твердые экскременты перед выделением отдают всю влагу стенками прямой кишки, как бы отжимаясь здесь досуха. Выдыхаемый воздух также успевает отдать всю влагу в густо оперенных ноздрях птицы. Наконец, пахучий экскрет из слепых кишок выделяется только при оставлении места ночлега утром, а в экстремальных обстоятельствах это выделение задерживается на сутки и более.
Длительность нахождения в подснежной камере регулируется каждой особью в зависимости от погодных условий, фотопериода, доступности кормов и своего физиологического состояния. При очень низких атмосферных температурах (от —30°С и ниже) такие виды, как Bonasa bonasia, B. umbellus, Lyrurus tetrix, могут ограничить всю свою активность двукратной или даже однократной кормежкой в сутки общей продолжительностью в 30—60 мин — все остальное время проводится в подснежной камере. На рис. 2 обобщены все данные, собранные мною к настоящему времени в природе о длительности пребывания тетеревиных птиц в подснежных камерах при тех или иных атмосферных температурах. Можно считать, что начиная с температуры —10 °С и ниже все тетеревиные птицы в середине зимы проводят в подснежных камерах (при наличии такой возможности) 60% времени суток и больше, причем чем севернее широта и больше длительность ночи, тем это время больше, но до определенных пределов. Например, у Lagopus lagopus на северо-востоке Азии общее время пребывания вне лунки (кормовая активность) во второй декаде декабря составила на 69° с. ш. 3.4 ч, а на 65° с. ш. — 3.7 ч в сутки (Андреев, 1982). При более высоких температурах время нахождения в подснежной камере быстро уменьшается как за счет увеличения времени кормежки, так и за счет отдыха на поверхности снега или на деревьях. Наконец, при температурах, близких к 0 °С и выше, тетеревиные птицы вообще не используют подснежных камер — в таких случаях вся нужда в термических убежищах отпадает. В результате получается, что тетеревиная птица, увеличивая или уменьшая длительность своего пребывания в подснежной камере, в известных пределах регулирует температуру окружающей ее внешней среды (табл. 1). Как показывают мои расчеты, при морозной, без оттепелей зиме эта среднесуточная температура составляет около —7 °С, причем птица способна поддерживать эту среднесуточную температуру при самых низких, по крайней мере до —50 °С атмосферных температурах.
Величина затрат на теплоотдачу во многом зависит от нижней критической температуры (Tнк) данного вида, т. е. от нижнего порога термонейтральной зоны, нахождение в которой не требует от птицы никаких затрат на терморегуляцию. Благодаря высоким теплоизоляционным качествам своего оперения и способности увеличивать их путем распушения перьев тетеревиные птицы имеют самую низкую Tнк среди изученных в этом отношении птиц, составляющую у L. lagopus, например, —6.5 °С (West, 1972; см. табл. 1), т. е. практически этот показатель приближается к температуре, поддерживаемой в подснежной камере, и находящаяся здесь птица пребывает в термонейтральной зоне. Экономия на терморегуляцию при использовании подснежной камеры составляет от 20 до 66 % от суточного бюджета энергии (табл. 1, 2).
До сих пор остается неясным, какое максимальное время может проводить птица под снегом, не покидая подснежной камеры. Ряд сообщений будто бы указывает на то, что при неблагоприятных погодных условиях (в сильный ветер при низких температурах) тетеревиные птицы способны оставаться под снегом по 1—2 суток (Снигиревский, 1946; Теплов, 1947а; Seiskari, 1962; Павлов, 1975). Расчеты по суточному бюджету энергии показывают, что для L. lagopus, например, при максимальной наполненности зоба и при наиболее высоком значении МЕС (коэффициента усвоения пищи), равного 0.45, находящейся в этом корме энергии хватит на 28 ч, после чего неизбежно начнется трата внутренних резервов птицы. Поэтому трудно предполагать, чтобы в природе тетеревиные птицы могли проводить в камерах больше, чем 1—1.5 сут подряд.
С регулярными ночевками тетеревиных птиц под снегом связано еще одно широко распространенное заблуждение, касающееся гибели их под снегом при обледенениях. Как было показано выше, подснежная камера — это прежде всего термическое убежище, становящееся совершенно ненужным в оттепели.
Более того, при оттепелях состояние снега таково, что делает невозможным зарывание в него птицы без того чтобы не замочить оперение. Поэтому теоретически имеется только одна вероятность оказаться под снегом в ледяном плену — для этого нужно, чтобы в течение одной ночи мороз сменился бы оттепелью, причем достаточно сильной, чтобы размочить поверхность снега, а затем — снова морозом, создавшим бы на поверхности снега толстую ледяную корку. Вероятность такого случая очень мала, но даже если он и произойдет, птица сразу же по повышению температуры в камере может установить начало оттепели и покинуть подснежное убежище. Мои наблюдения показали, что тетеревиная птица без колебания оставляет подснежную камеру в любой момент ночи как при опасности, так и по другим причинам, причем без труда находит себе новое место для ночлега. Именно этот момент упускают из вида авторы, отстаивая старые представления о гибели птиц «в ледяном плену» (Формозов, 1976; Киселев, 1978, и др.). Все свидетельства на этот счет базируются на фактах нахождения мертвых тетеревиных птиц без видимых следов повреждения, иногда вытаивающих из-под снега весной. О находках такого рода неоднократно сообщали охотники и егери, связывавшие причину гибели птиц с ледяным настом. Такую же находку сделал и А. Н. Формозов, причем во всех случаях погибшие птицы находились в спокойных «спящих» позах, да еще с кормом в зобах, что никак не вяжется с гибелью от голода при отчаянных попытках разломать ледяную корку. Причины гибели таких птиц до сих пор не установлены, но у нас нет никаких оснований считать ею ледяной наст.
Оледенения, следующие за оттепелями, действительно могут вызвать гибель тетеревиных птиц, но совсем по-иному. Известны случаи, когда после длительной сильной оттепели наступали морозы, одевавшие всю древесно-кустарниковую растительность и поверхность снега ледяной коркой. Резко возросшие затруднения с добычей корма и невозможность зарыться под снег вызывали в конце концов массовую гибель тетеревиных птиц от истощения, но она наступала лишь при длительном сохранении таких условий, не менее 4 недель (McGowan, 1969). Столь длительные обледенения — явление весьма редкое и не являющееся, постоянно действующим фактором среды. Более же короткие обледенения, длящиеся менее недели, не наносят тетеревиным птицам заметного ущерба.
Помимо термического, подснежная камера является также и маскирующим убежищем, предохраняющим тетеревиных птиц от нападения с воздуха, т. е. от хищных птиц, как дневных, так и ночных. Это особенно существенно для мелких тетеревиных птиц, так как устраняют угрозу нападения ночью сов. В то же время пребывание под снегом не спасает птиц от нападения хищных зверей, прежде всего куньих, лисицы, песца, енотовидной собаки, росомахи и волка. Куница, например, как и лисица, может почуять глухарей, находящихся под снегом, с большого расстояния и довольно точно рассчитать скорость и направление атаки (Семенов-Тян-Шанский, 1959; Киселев, 1978). Мне неоднократно приходилось находить следы успешной охоты на тетеревиных птиц в лунках лисицы и даже волка. Примечательно, что цепочки лисьих следов зимой особенно часто идут по местам, наиболее благоприятным для ночевки полевого тетерева.
Насколько велики неудобства, связанные для тетеревиных птиц с передвижением по снегу, доказывают расчеты Андреева (1980), показывающие, что при самых неблагоприятных условиях птицы затрачивают на хождение по рыхлому снегу не более 3—3.8 % от суточного бюджета энергии. Даже эти низкие показатели кажутся мне завышенными по крайней мере вдвое, ибо при своих расчетах А. В. Андреев исходил из того, что птица с каждым шагом поднимает свое тело на высоту, равную глубине проваливания. Мои наблюдения показывают, что центр тяжести птицы при ходьбе по рыхлому снегу перемещается почти горизонтально, а лапы, подгибаясь, протаскиваются сквозь снег и погружаются с каждым шагом до появления точки опоры. Положение этих точек опоры — спрессованного под лапой снега — относительно поверхности одно и то же. Таким образом, к энергетической стоимости простого хождения следует прибавить только ту энергию, которая необходима для протаскивания лапы сквозь снег. Точных измерений этих усилий не делалось, но они явно значительно меньше тех, что необходимы для подъема всего тела вверх, на глубину проваливания. Поэтому предварительные расчеты затрат на хождение по рыхлому снегу у таких многоходящих видов, как Lagopus lagopus или Tetrao urogalloides, оцениваются как не превышающие 1.5 % от суточного бюджета энергии. Следует также иметь в виду, что, за исключением видов рода Lagopus, практически все тетеревиные птицы в середине зимы могут обходиться без хождения по снегу и если они это и делают, то не всегда ясно, что именно побудило их к этому.
Пребывание в подснежной камере влечет за собой еще один крупный энергетический выигрыш — снижение двигательной активности. Сравнительная легкость добывания корма, имеющегося, как правило, в достаточном количестве, а также большая емкость зоба, вмещающего по сути дела весь объем суточного рациона, позволяют затрачивать на сбор этого рациона минимальное (от 1 до 3 ч) время, освобождая остальную часть суток для отдыха и сна. Хронометраж зимней активности тетеревиных птиц, выполненный как мною, так и другими авторами (Андреев, 1980, 1982), показывает, что на двигательную активность в середине зимы рябчик, например, затрачивает всего 3 ч в сутки, в том числе на полет не более 2—3 мин. Сходные цифры характерны и для остальных видов (табл. 2). Это резкое снижение активности зимой, вместе с устройством подснежных камер, является исключительно важной поведенческой адаптацией, позволяющей эффективно реализовать все возможности, даваемые птице функциональными способностями ее пищеварительного тракта, прекрасными термоизоляционными свойствами оперения, а также такими морфологическими структурами, как рамфотека и роговые выросты на пальцах.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
 Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты).
Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты). Суточный бюджет энергии и времени тетеревиных птиц.
Суточный бюджет энергии и времени тетеревиных птиц.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Гнездование тетеревиных птиц.
Гнездование тетеревиных птиц. Брачный ритуал тетеревиных птиц - токование.
Брачный ритуал тетеревиных птиц - токование. Три дня с дрофой наедине.
Три дня с дрофой наедине. Птица стенолаз.
Птица стенолаз. Птицы щурки.
Птицы щурки.
