Размножение тетеревиных птиц. Характер брачных отношений.
У тетеревиных птиц наблюдается полная гамма переходов от строгой моногамии до ярко выраженных полигамных отношений при групповом характере спаривания на так называемых токах. По характеру взаимоотношений полов семейство можно разделить на 3 группы (Hjorth, 1970; Wiley, 1974). Первую группу (табл. 1) образуют виды, для которых характерно сочетание полигамии с одиночным токованием самцов на специальных, защищаемых ими участках. Защита участков выражена только в период токования и быстро исчезает после того, как самки приступают к насиживанию. На своем участке самец выполняет токовой ритуал, изгоняет посторонних самцов и спаривается со всеми привлекаемыми им самками. Размеры участка обычно значительны (0.4—5 га), и самки, спарившиеся с хозяином участка, нередко здесь гнездятся. Однако считать этот участок гнездовым нельзя, поскольку одна или несколько самок, избравших данного самца, могут гнездиться как в пределах его участка, так и вне его, и территориального антагонизма между самками нет. На токовом участке самцом используется не вся его территория, здесь имеется центр активности с несколькими точками, где самец токует особенно часто. Обычно токовые участки нескольких самцов располагаются в непосредственной близости один от другого, и группы таких участков чередуются с ненаселенной данным видом территорией.
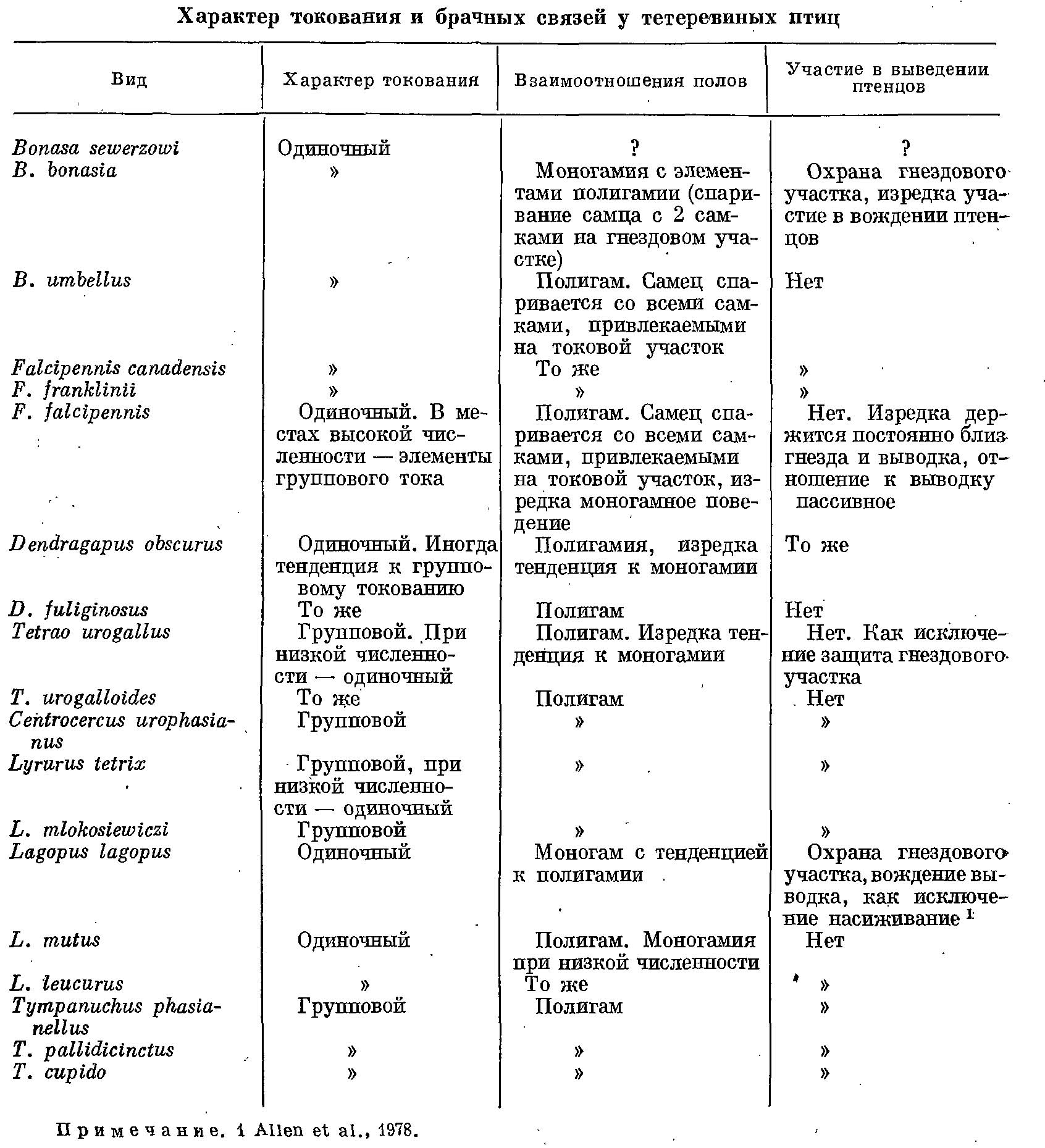
Таблица 1. Характер токования и брачных связей у тетеревиных птиц.
Вторая группа тоже полигамная, но для самцов характерно групповое токование. Место токования — «токовище» — нередко становится репродуктивным центром популяции. Самцы собираются на эти токовища ежедневно в определенные часы, каждый из них имеет свой определенный участок, на котором он токует, спаривается и который охраняет от вторжения соперников. Обычно групповой ток имеет сложную иерархическую структуру из трех ступеней. Наиболее активные самцы занимают самые важные, ключевые участки, менее активные — окраинные и вообще менее удобные места, и, наконец, молодые самцы нередко совсем не имеют своих участков. Ранг иерархии зависит в основном от возраста птицы, и наиболее активные самцы, занимающие вершину иерархической пирамиды и оплодотворяющие большинство самок, обычно имеют возраст от трех и более лет. Эта идеальная схема редко наблюдается в чистом виде и часто нарушается. Например, у таких видов, как Tetrao urogallus или Lyrurus tetrix (мои наблюдения), на одном и том же токовище картина даже в течение одного утра может резко меняться: то весь ток неожиданно перемещается, на несколько десятков, а то и сотню метров, то вдруг перестают соблюдаться границы участков. В ряде случаев никакой возрастной иерархии нет, и молодые самцы с равным успехом участвуют в токовании (Tympanuchus phasianellus; Hjorth, 1970). У некоторых видов (Tetrao urogallus, Lyrurus tetrix, Centrocercus urophasianus) обширный ток, собирающий до сотни и более самцов, дробится на более мелкие группы, в каждой из которых токование идет, как на отдельном небольшом токовище (Scott, 1942). Пространственная структура тока может сильно варьировать, нередко даже в соседних микропопуляциях один и тот же вид может иметь то диффузный и обширный ток с большими индивидуальными участками, то очень концентрированный, где индивидуальные участки в центре занимают не более нескольких десятков квадратных метров. На индивидуальных участках имеются определенные места (в том числе и определенные деревья), где самцы токуют особенно часто. Как мы видим, территориальное поведение самцов в этой группе ограничивается как в пространстве (только на токовище), так и во времени (определенное время суток в брачный сезон). Самцы этой группы тоже не принимают участия в выводе птенцов.
Третья группа представлена только двумя видами — Bonasa bonasia, Lagopus lagopus. Это моногамы, у которых участок самца является и токовым и гнездовым; здесь может гнездиться только одна самка, а пары могут образовываться даже с осени. Самец принимает участие в выводе птенцов, хотя степень этого участия различна.

Тетерев-косач (Lyrurus tetrix) спаривается с самкой.
Главные отличия первых двух групп от последней — использование самцом участка для токования и более позднее вступление самцов в размножение по сравнению с самками (обычно в возрасте 2 лет). В первых двух группах молодые возрасте менее года самцы обычно не имеют своих токовых участков и не участвуют в размножении, хотя многие из них достигают половой зрелости (Немцев и др., 1973; Wiley, 1973; Кутовая, 1976). Самцы-моногамы становятся половозрелыми, как и самки, на следующую весну, хотя успешность их размножения по сравнению со старшими бывает меньшей (Jenkins et al., 1963; Willey, 1974). Наблюдается и более позднее (до 24 дней) гнездование молодых самок (Bonasa umbellus; Porat, Vohns, 1972).
Разграничение этих групп нерезкое. Некоторые авторы (Hjorth, 1970; Wiley, 1974) причисляют Lagopus mutus и L. leucurus к группе моногамов, хотя по характеру связи полов и территориальности самок они скорее относятся к первой группе. У Dendragapus obscurus, явно относящегося к первой группе, самка иногда гнездится на токовом участке самца и тот остается с ней весь период насиживания. Подобные случаи бывают, по-видимому, и у Falcipennis falcipennis. Даже у такого резко выраженного полигама, как Tetrao urogallus, известны случаи, когда самцы охраняли насиживающую самку (Bannerman, 1963). У L. tetrix, для которого столь характерно групповое токование, в ряде случаев самцы токуют в одиночку: такие самцы встречаются даже при нормальной плотности популяции (Hjorth, 1970; мои данные). Эти самцы ведут себя, как самцы первой группы, но держатся на токовых участках не постоянно, а только в определенные часы, как и на коллективных токовищах. Такое же явление было обнаружено мною и у T. urogallus.
У фазановых птиц также наблюдается заметное разнообразие во взаимоотношениях полов. Однако характер этого разнообразия здесь совсем иной. Во-первых, среди Phasianidae нет группового токования — этот тип полигамии у тетеревиных птиц по сравнению с фазановыми совершенно новый. Во-вторых, для большинства видов характерно круглогодичное обитание стайками, даже в брачное время. По моим наблюдениям, у ряда видов (Perdicula asiatica, Alectoris kakelik, Tetraogallus spp.) с началом брачного сезона в такой стайке, ведущей полукочевой образ жизни и имеющей самые различные соотношения возрастных и половых групп, возникает заметное оживление. Самцы начинают токовать прямо в стае и по мере развития брачного состояния то одна, то другая спонтанно образующиеся пары отделяются от общей группы и уединяются для гнездования. Образование пар определяется степенью готовности отдельных особей к размножению. У небольших по размерам фазановых птиц половозрелость достигается самцами и самками на первом году жизни, и они участвуют в размножении наряду со старыми птицами, но в более поздние сроки. Это сильно растягивает как сезон размножения, так и распад стай на отдельные пары. Нередко такая стая так и не распадается до конца — некоторые птицы остаются, к ним присоединяются самки, потерявшие кладки, и самцы, покидающие выводки, а затем и новое поколение. Известны даже случаи, полиандрии, когда самка оставляет на попечение самца подрастающих птенцов, а сама, вновь спариваясь, начинает новую кладку (Odontophorinae, Bent, 1932; Alectoris, Johnsgard, 1973; Грачев, 1978). Однако моногамия преобладает, самец участвует в защите гнезда, выводка и даже в инкубации. При такой социальной структуре нет ни одиночного, ни группового территориального токования, несложный токовой ритуал совершается и в стае, состоящей из особей различных по полу, возрасту и степени готовности к размножению и в обособившейся паре. Такие ситуации у тетеревиных птиц отсутствуют, но как своеобразный атавизм проявляются иногда на ранних стадиях развития весеннего возбуждения в смешанных стаях (иногда состоящих и из одних самцов). В тихую солнечную погоду на снежном насте среди самцов спонтанно возникает брачное возбуждение — они начинают выполнять отдельные элементы токового ритуала и даже преследовать самок, на которых возбуждение не распространяется (Т. urogallus, L. tetrix, мои данные; В. bonasia, Донауров, 1947; L. lagopus, Михеев, 1948). Для фазановых птиц обычна и ситуация, характерная для первой группы тетеревиных. Самцы токуют в одиночку на своих участках и спариваются с привлекаемыми ими самками (Phasianus, Lophophorus, Tragopan и др.).
Вполне вероятно, что у отдаленных предков тетеревиных существовали взаимоотношения, подобные примитивной моногамии многих рецентных видов подсемейств Perdicinae и Odontophorinae. Но уже на первых этапах становления предковой формы могла существовать полигамия при одиночном характере токования. Такая система кажется наиболее простой и из нее легко выводятся более специализированные системы взаимоотношений — как групповое токование, так и строгая моногамия.
Полигамия, развитая у тетеревиных птиц вплоть до самых крайних форм, несомненно дает определенное селективное преимущество популяции и виду в целом — без этого она не могла бы развиться. Однако сам процесс возникновения и развития группового токования, его адаптивная и эволюционная значимость до сих пор не поняты до конца, о чем свидетельствует непрекращающаяся полемика вокруг этих вопросов. Здесь предполагается и отстранение от участия в размножении молодых самцов, что нарушает соотношения полов в пользу самок, но способствует выживаемости самцов до возраста, когда их репродуктивный потенциал максимален; и возрастной кросс; и снижение потерь от хищников; и изобилие пищи, делающее ненужной гнездовую территорию и освобождающее самца от заботы о потомстве; и максимальная эффективность оплодотворения самок, а также ряд других предположений, причем все они исходят из несомненного адаптивного преимущества группового тока (Armstrong, 1949, 1955; Amadon, 1959; Selander, 1965, 1966; Чельцов-Бебутов, 1965; Short, 1970; Wittenberger, 1978, и др.).
Наиболее интересна, на мой взгляд, гипотеза Вилея (Wiley, 1974), который обратил внимание на то, что у всех (за исключением Tympanuchus phasianellus) полигамных видов тетеревиных самцы созревают позже самок, причем с увеличением размеров и ростом полового диморфизма эта разница в скорости созревания полов возрастает. Это явление, известное у птиц и млекопитающих под термином «сексуальный биматуризм», нарушая реальное соотношение полов в группе половозрелых птиц, способствует развитию полигамии. Причина задержки развития самцов, как ее трактует Вилей, вполне возможна: чем позднее вступает самец в размножение, тем больше у него шансов дожить да более взрослого состояния, когда его репродуктивная способность становится максимальной. Однако эта гипотеза, как и все остальные, сталкивается с одной неразрешимой проблемой. Если адаптивные преимущества полигамии и группового тока столь очевидны, то почему же они не развились в равной степени у всех видов семейства, весьма близких по своей биологии? Почему даже в пределах одного вида мы наблюдаем обратимые переходы от одиночного к групповому токованию и наоборот? Кажется несомненным наличие каких-то причин, нам пока неизвестных, которые способствуют развитию группового токования в одних случаях и одиночного — в других. Например, влияние ландшафта, вроде бы, сказывается на характере токования — наблюдается несомненная тенденция к групповому токованию у видов открытых ландшафтов и к одиночному — в лесах (Hjorth, 1970; Wiley, 1974). Однако одиночное токование белых куропаток, обитающих в открытых ландшафтах, и групповое лесных обитателей — глухарей — явно противоречит такой тенденции. Более того, из двух наиболее строгих моногамов семейства один является обитателем леса (В. bonasia), другой — открытых пространств (L. lagopus). Различные типы брачных взаимоотношений у видов, живущих бок о бок в одних и тех же экосистемах (Т. urogalus и В. bonasia, L. tetrix и L. lagopus), говорит о том же — невозможности связать их только с современными экологическими условиями. Столь же трудно искать причины развития таких взаимоотношений в соотношении полов в популяциях. Детальные полевые исследования показали, что соотношение самцов и самок в одних и тех же популяциях заметно меняется из года в год (Киселев, 1971 и др.). Эти обстоятельства заставляют обратить особое внимание на наследственные факторы. Поскольку наблюдается определенное сходство в типах брачных связей в пределах рода или в крайнем случае подрода, то можно признать наследственные факторы более существенными, нежели современная экологическая обстановка. В качестве рабочей гипотезы можно предположить, что характер биологии вида так же, как и тип ландшафта действительно оказали сильное влияние на формирование брачных взаимоотношений, но это происходило только на первых этапах становления семейства, в процессе адаптивной радиации. Впоследствии типы брачных связей были закреплены в наследственности и подвергались в дальнейшем сравнительно малым модификациям, хотя отдельные виды, у которых выработалось групповое токование, сохранили возможность возврата к исходному состоянию (L. tetrix,, Т. urogallus).
Пути развития группового токования можно себе представить следующим образом. Первоначальной, на мой взгляд, является система, характерная для видов 1-й группы (табл. 1). Самцы этих видов токуют в одиночку, но групповое расположение токовых участков, их тесное соседство приводит к возбуждающему действию токующих самцов друг на друга, а молодые «безземельные» самцы привлекаются к местам скопления территориальных самцов (Blackford, 1959; Bendell, Elliot, 1966; Gullion, 1967, и др.). В этом можно усмотреть начало процесса, который ведет к возникновению группового токования. Скопления территориальных самцов в определенных местах становятся все больше, участки, занимаемые ими, все меньше, возбуждающее влияние друг на друга — все ощутимее. Участки теряют всякое значение для гнездования и кормежки и служат почти исключительно местом токования и спаривания. Принцип остается тем же — самец привлекает, а самка выбирает, но коллективное токование резко повышает возбудимость обоих полов и способствует большей синхронизации полового цикла в популяции. Каждый достаточно крупный коллективный ток становится репродуктивным центром микропопуляции и обеспечивает высокую синхронность размножения.
Развитие брачных отношений может идти и в противоположном направлении — к моногамии. Территориальные самцы при значительной плотности популяции все более активно защищают свои участки, размеры которых достаточны для гнездования самки. Охранные и отвлекающие функции, которые выполняет самец на таком участке, должны снижать смертность самок и тем самым повышать концентрацию «территориальных» генов в генофонде конкретной популяции. Во всяком случае стабильность типа брачных связей в пределах вида в настоящее время отражает уже несомненное действие гомеостаза, определяемого устойчивостью сбалансированных регуляторных систем.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
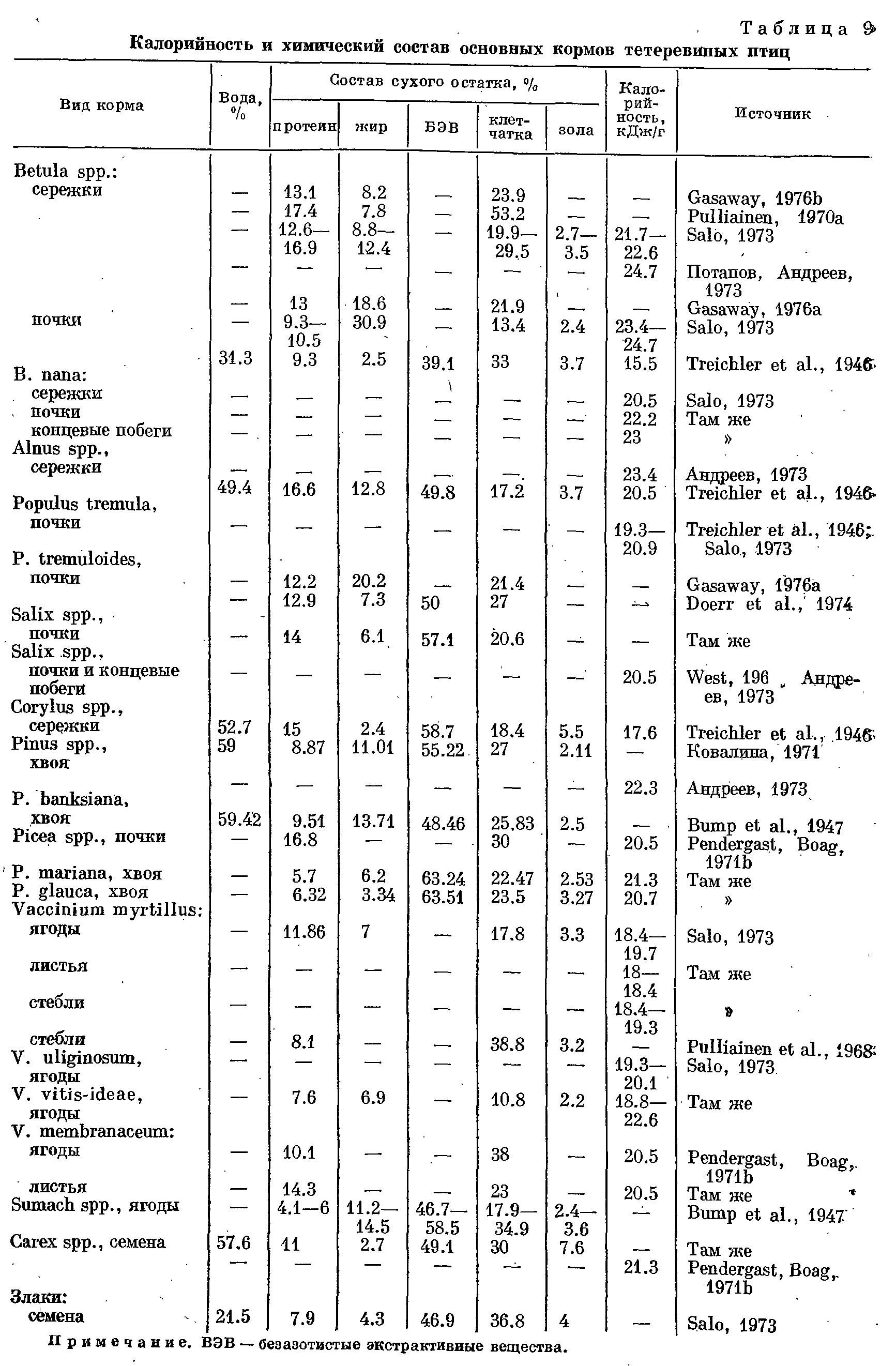 Факторы, определяющие сезонное изменение питания тетеревиных птиц.
Факторы, определяющие сезонное изменение питания тетеревиных птиц. Вторично-половые признаки тетеревиных птиц. Половой диморфизм.
Вторично-половые признаки тетеревиных птиц. Половой диморфизм.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Питание тетеревиных птиц.
Питание тетеревиных птиц. Особенности мест обитания тетеревиных птиц.
Особенности мест обитания тетеревиных птиц. Ареал тетеревиных птиц, их распространение и образ жизни.
Ареал тетеревиных птиц, их распространение и образ жизни. Фотоохота на серую цаплю.
Фотоохота на серую цаплю. Слепой отдел кишечника тетеревиных.
Слепой отдел кишечника тетеревиных.
