Вторично-половые признаки тетеревиных птиц. Половой диморфизм.
Вторично-половые структуры у тетеревиных птиц развиты незначительно. Это прежде всего «брови» — особые кожные образования над орбитами, затем так называемые шейные мешки, украшающие перья и с некоторыми оговорками хвост, в размерах, форме и цвете которого половой диморфизм проявляется особенно ярко. Однако развитие всех этих структур не всегда коррелирует с развитием полового диморфизма и степенью полигамии.
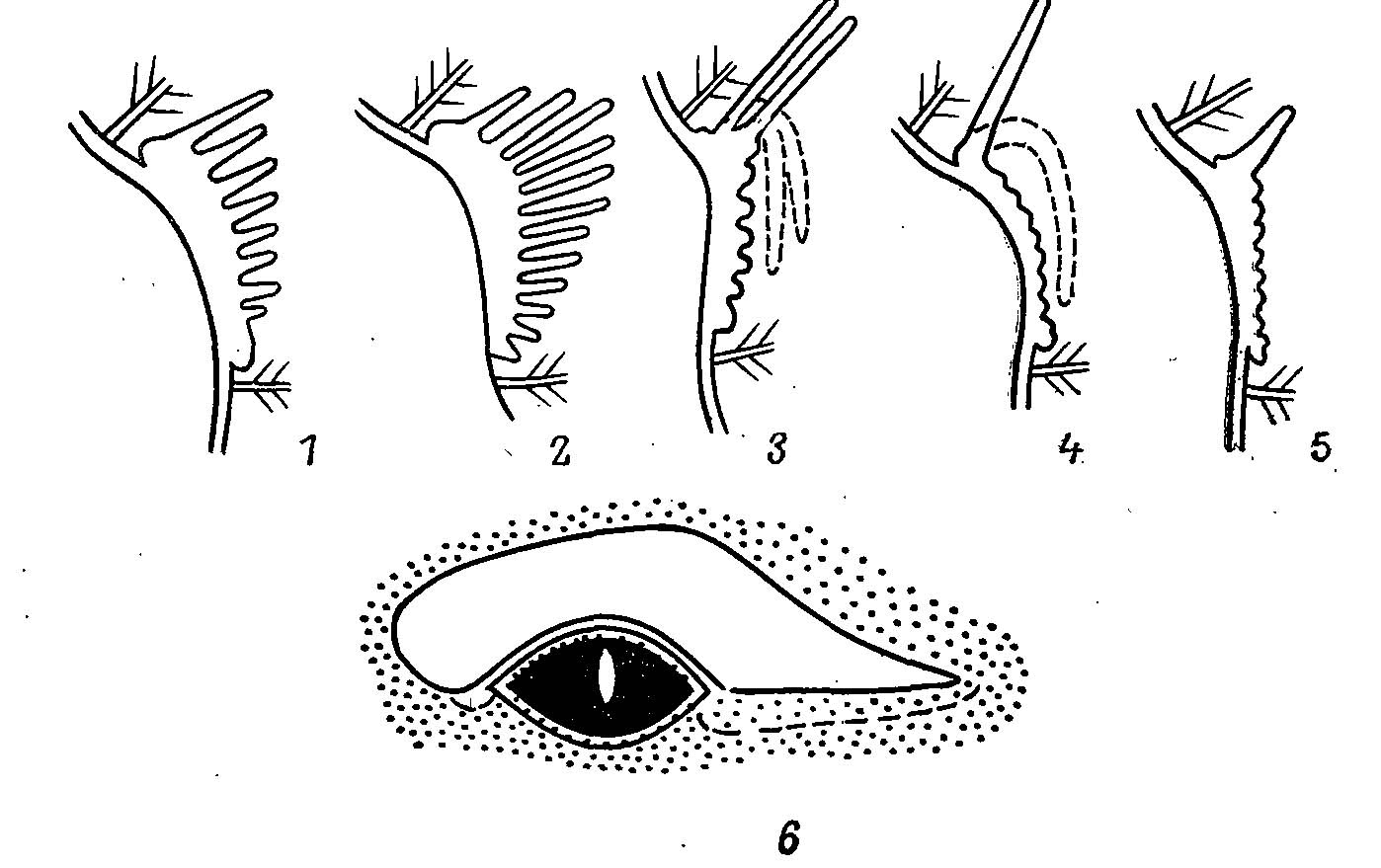
Рис. 1. Строение «брови» у тетеревиных птиц (поперечные срезы). 1 — Bonasa, Falcipennis, Dendragapus, Tetrao; 2 — Lyrurus tetrix; 3 — L. mlokosiewiczi; 4 — Lagopus; 5 — Tympanuchus, Centrocercus (прерывистая линия — положение лопасти в спокойном состоянии, 3, 4); 6 — распространение красного цвета на «брови» глухаря (сплошная линия — «бровь») и соседних участках кожи (прерывистая линия). Точки — места прикрепления перьев.
У всех тетеревиных птиц над глазом расположена «бровь» красного цвета — участок кожи, лишенный перьев, где развита рыхлая, сильно васкуляризированная ткань, покрытая бугорками или палочкообразными выростами и пронизанная густой сетью кровеносных сосудов. В большинстве случаев по верхнему краю идет невысокий гребень (рис. 1). Большую часть года «бровь» не функционирует и представляет собою узкую полоску, нередко скрытую соседними перьями. Резкое увеличение «брови» за счет набухания и выпячивания бугорков и выростов происходит у самцов в период спаривания. Цвет их в моменты полового возбуждения становится ярко-красным, что вызывается, помимо пигментов ксантофила и каротина (Lonnberg, 1930), переполнением тканей кровью. В зависимости от притока крови цвет «бровей» может меняться у одной и той же особи от желто-красного до ярко-алого в течение нескольких минут. Противоречивые описания цвета «бровей» некоторых видов в литературе вызваны именно этим обстоятельством (Lyrurus mlokosiewiczi, Lorenz, 1884).
У видов рода Tympanuchus цвет «бровей» не столь ярок, как у остальных, вследствие того, что главным цветовым раздражителем у них становятся шейные мешки. Обратная зависимость между размерами «бровей» и шейных мешков, подмеченная Хьортом (Hjorth, 1970), справедлива только для этого рода и Centrocercus urophasianus. У рода Dendragapus оба эти образования развиты достаточно хорошо. По окраске «бровей» полынный тетерев стоит особняком: цвет их описан как «оливково-зеленый» (Hjorth, 1970).
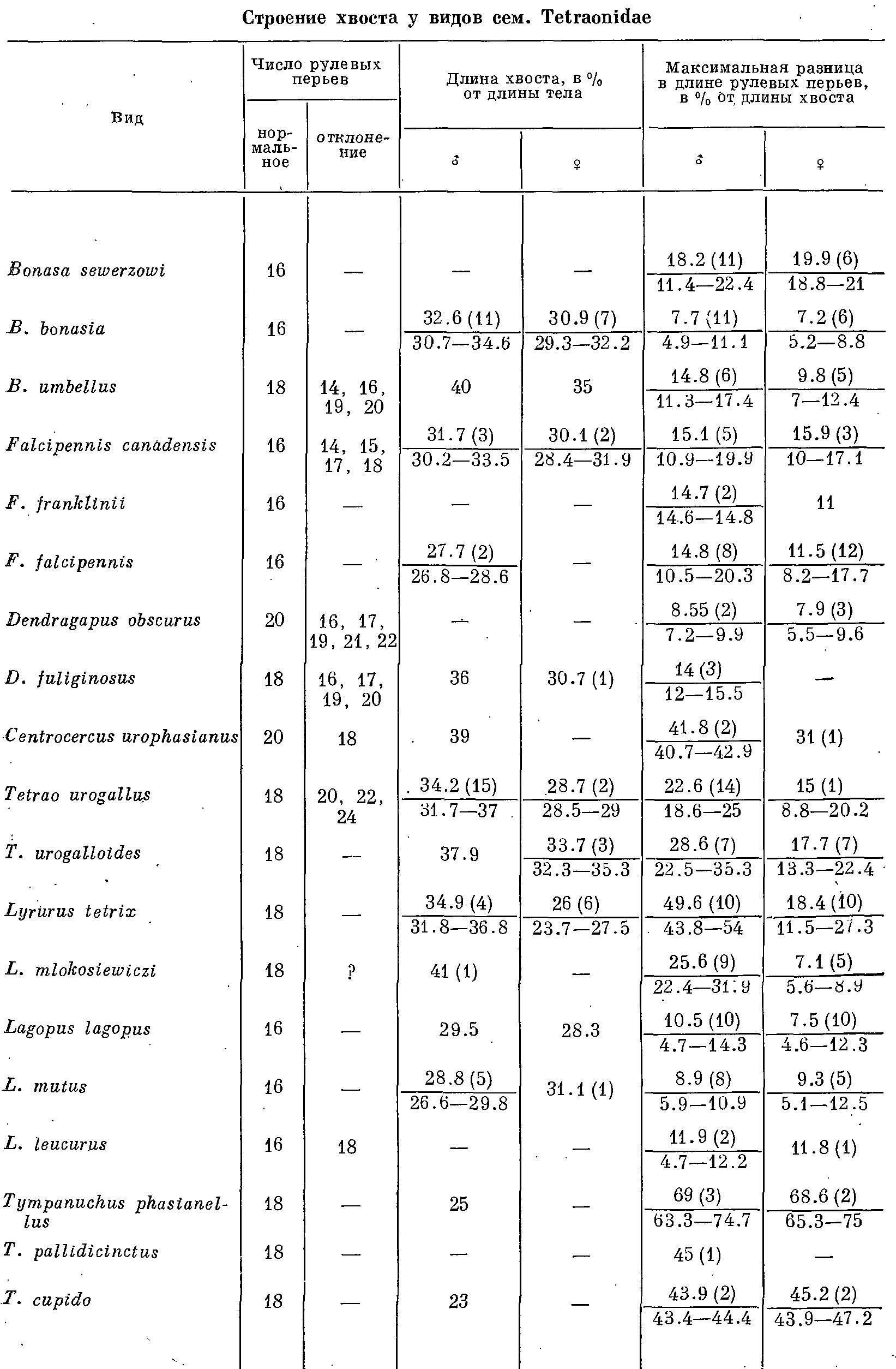
Таблица 1. Строение хвоста у видов сем. Tetraonidae.
Способность птиц резко менять объем «бровей» в разгар брачного сезона является важной адаптацией к токованию при низких температурах воздуха. У наиболее северных видов (род Lagopus) лопасть «брови» во время токования достигает значительных размеров, но в остальное время, на отдыхе или кормежке, размеры ее заметно уменьшаются, она свисает вниз, складываясь вдвое, и полностью скрывается соседними перьями. Это позволяет птице избегать ее обморожения и уменьшает потерю тепла. Регуляция размера «бровей» позволяет тетеревиным птицам сохранять эти яркие вторично-половые структуры в условиях холодного климата. Обилие яркоокрашенных участков и выростов голой кожи на голове, которое наблюдается у многих видов фазановых птиц, не наносит ущерба птице только в теплом климате.
«Брови» у тетеревиных птиц бывают двух типов. У одних вся площадь брови покрыта высокими пальцевидными выростами, часть из которых срастается у основания. Относительная высота этих выростов у разных видов неодинакова и постепенно возрастает в ряду В. bonasia, Falcipennis spp., Dendragapus spp., Tetrao spp., L. tetrix. У всех, за исключением последнего (рис. 1), максимальной высоты достигает верхний ряд выростов. У других видов по верхнему краю развивается широкая лопасть, нередко равная по площади самой «брови», с гребенчатым краем, тогда как рельеф на самой «брови» выражен невысокими бугорками. Этот тип свойствен видам родов Lagopus, Tympanuchus и Centrocercus. Примечательно строение «брови» у L. mlokosiewiczi (рис. 1) — явно промежуточный случай, показывающий путь развития «брови» такого типа. Оба типа вполне могли развиться из того неспециализированного состояния, которое в настоящее время характерно для В. umbellus. В первом случае выросты на «брови» будут увеличиваться на всей ее поверхности, особенно в верхней части, а во втором — максимально разрастается только самый верхний ряд выростов, основания которых срастаются и образуют сплошную лопасть.
Если относительная длина выростов максимальна у L. tetrix, то по абсолютной площади «брови» на первом месте стоит Т. urogallus. «Бровь» у него начинается впереди глаза и тянется далеко назад, а пигментация захватывает и прилежащую часть глазной птерилии — как бы намек на возможность образования вокруг глаза сплошного кольца голой окрашенной кожи, как у некоторых фазановых птиц (рис. 1).
Шейные мешки — это особые структуры, в образовании которых участвуют кожа в области шейных аптерий, подкожная мускулатура и расширенный участок эзофагуса перед зобом. Эти структуры описаны у Tympanuchus cupido (Gross, 1928) и С. urophasianus (Honess, Allred, 1942). При токовании сильный сфинктер перекрывает вход из эзофагуса в зоб, расширенный участок эзофагуса под действием поступающего из легких воздуха сильно раздувается и выпячивает наружу эластичные участки шейных аптерий. Последние, несущие в это время яркую окраску, вздуваются двумя красными или оранжевыми пузырями по бокам шеи. Роль этих образований как резонаторов вполне очевидна, поскольку в моменты ее максимального расширения птица обычно издает различного рода звуки (исключение С. urophasianus), но прямой связи между силой звука и степенью развития шейных мешков нет. Например, песня L. tetrix, не имеющего шейных мешков, слышна до 3 км, тогда как у такого же по размерам Dendragapus fuliginosus слышимость пения не превышает 600 м. Более того, у одинаковых по размерам видов дымчатых тетеревов шейные мешки развиты также одинаково, но слышимость вокализаций в одном случае — 100 м (Mussehl, 1963), во втором — около 600 м (Bendell, 1955; Hjorth, 1970). Здесь, видимо, более важна функция шейных мешков как визуальных раздражителей. Первоначально расширение эзофагуса явно способствовало усилению звука: у F. falcipennis и L. tetrix эзофагус сильно расширяется при вокализациях, но не настолько, чтобы выпячивать кожу шейных аптерий, хотя у последнего вида в отдельные моменты токования она иногда бывает видна. При дальнейшем увеличении расширяемой части эзофагуса неизбежно происходило регулярное экспонирование вздувающихся участков шейных аптерий, где впоследствии развилась пигментация. Только у С. urophasianus формирование шейных мешков пошло по более сложному пути. Устройство и функциональные особенности этих мешков указывают на независимое их развитие у Centrocercus, Dendragapus и Tympanuchus — интересный случай параллельного развития сравнительно сложной структуры на одной и той же морфологической основе.
Украшающие перья, заметно отличаясь от остальных, имеют значение только в брачный сезон. Это перья «воротничка» В. umbellus, «уши» Т. pallidicinctus и Т. cupido и «вымпеловые» перья на голове С. urophasianus. Перья такого типа расположены только на голове, шее и груди и, имея различные форму и окраску, выполняют одну функцию, — поднимаясь, они заметно преображают контур птицы и даже ее окраску. Особенно необычны перья, сплошь покрывающие часть шеи и груди у Centrocercus. Их своеобразная форма — резкое заострение толстого и короткого стержня — придает необходимую жесткость перу, главной функцией которого становится продуцирование своеобразного звука: при противоположно направленных движениях расширенной шеи и прижатых к ней крыльев нижняя поверхность крыла скребет по перьям шеи, как ноготь по гребенке (Brooks, 1930; Hjorth, 1970).
Активное участие хвоста в брачном ритуале всех видов семейства обусловило развитие полового диморфизма как в окраске, так в форме и размерах рулевых перьев. У самок окраска хвоста проста и однообразна — это поперечно-полосатый рисунок с чередованием крупных черно-бурых и светло-желтых полос. Форма рулевых перьев большей частью нормальная, а их длина заметно короче, чем у самцов.
По мере возрастания роли хвоста в брачном ритуале наблюдаются определенные изменения в величине, форме и окраске рулевых перьев, а также заметное увеличение их числа до 18—20. Увеличивается общая длина хвоста (табл. 1), причем нередко возрастает разница в длине рулевых перьев — длина их уменьшается от центра к краям и только у рода Lyrurus картина обратная. В ряде случаев вершина пера становится прямо срезанной, приобретая своеобразный скобковидный абрис (В. bonasia, В. sewerzowi, Lagopus spp., Т. phasianellus), а ширина пера заметно увеличивается. У других видов прямо срезанная вершина пера отгибается в сторону (Lyrurus spp.). Наконец, у Centrocercus перья хвоста заострены. Несколько особняком стоит Т. phasianellus, где четко различаются длинная, со слегка закругленными вершинами центральная пара и укороченные изогнутые остальные рулевые перья. Функция хвоста в брачном ритуале у этого вида наиболее сложная.
Изменение окраски по мере возрастания роли хвоста в токовании идет (за одним исключением) по линии усиления пигментации и развития равномерной черной окраски. В наиболее примитивном состоянии хвост имеет грубый поперочно-полосатый рисунок (Bonasa spp.). Усиление пигментации и развитие сплошной окраски идет от центра к краям хвоста — на крайних перьях остатки поперечно-полосатого рисунка удерживаются дольше всего. Белая вершина, наоборот, дольше всего удерживается на центральной паре. Так, у Т. urogalus, где хвост взрослых самцов не имеет и намека на белую вершину, остатки ее видны на центральных перьях у годовалых птиц. Эта общая тенденция имеет свои особенности у разных видов, среди которых наиболее общее явление — иная окраска центральной пары. Последняя бывает окрашена иначе, чем остальные, под цвет поясницы и выполняет маскировочную функцию, скрывая на сложенном хвосте отнюдь не криптическую окраску остальных перьев (В. bonasia, B. sewerzowi, Lagopus spp., Т. phasianellus). Критическая роль центральной пары особенно ярко проявляется у видов рода Lagopus — она даже линяет в отличие от остальных дважды в год. Белая зимой и пестрая летом, она скрывает сплошную черную окраску сложенного хвоста. При дальнейшем усилении пигментации центральная пара приобретает ту же окраску, что и остальные, но первоначальная ситуация может сохраняться в 1-м взрослом наряде (F. falcipennis).
Развитие сплошной черной окраски хвоста обычно сопряжено с образованием контрастного рисунка из белых вершин верхних и нижних кроющих перьев хвоста при темной окраске остальной части пера. На черном фоне раскрытого хвоста эти вершины образуют довольно правильные полуокружности — контрастный видоспецифичный рисунок, особенно резко заметный в сумерках. Эта тенденция максимально выражена у Т. urogalloides. У Т. urogallus она модифицирована развитием на рулевых перьях белых полей, в связи с чем произошла мастичная редукция белых вершин подхвостья и особенно надхвостья. Другая модификация — развитие сплошь белого подхвостья (L. tetrix, Т. cupido), резко наделяющегося на черном фоне раскрытого хвоста.
Все вторично-половые признаки самцов в какой-то степени выражены и у самок. Только окраска хвоста остается специфично самочьей, но его форма у самок является либо слабым отражением формы хвоста самцов (Lyrurus spp.), либо почти неотличима от нее, несмотря на полное отсутствие соответствующей функции (Tympanuchus spp., Centrocercus). «Брови» имеются у всех самок, они даже несколько увеличиваются весной, но никогда не достигают сколько-нибудь заметной величины. Шейные мешки у самок не развиты, но у С. urophasianus специфичные аптерии на фронтальной части шеи у самок такие же, как и у годовалых самцов. Украшающие перья у самок тоже есть, но их размеры значительно меньше. Только «вымпеловые» перья полынного тетерева свойственны исключительно самцам.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
 Размножение тетеревиных птиц. Характер брачных отношений.
Размножение тетеревиных птиц. Характер брачных отношений. Малые подорлики.
Малые подорлики.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Факторы, определяющие сезонное изменение питания тетеревиных птиц.
Факторы, определяющие сезонное изменение питания тетеревиных птиц. Питание тетеревиных птиц.
Питание тетеревиных птиц. Особенности мест обитания тетеревиных птиц.
Особенности мест обитания тетеревиных птиц. Ареал тетеревиных птиц, их распространение и образ жизни.
Ареал тетеревиных птиц, их распространение и образ жизни. Фотоохота на серую цаплю.
Фотоохота на серую цаплю.
