Суточный бюджет энергии и времени тетеревиных птиц.
Особенности зимней жизни тетеревиных птиц позволили мне в свое время разработать методику, впервые давшую возможность определить суточный бюджет энергии птицы в природе (Потапов, Андреев, 1973). Впоследствии эта методика была значительно усовершенствована (Потапов, 1974, 1982; Андреев 1975б, 1980, 1982; Потапов, Андреев, 1982). Она базируется на выяснении основных компонентов, позволяющих вычислить ежедневное потребление энергии. Компоненты эти следующие.

Рябчики.
1. Общее количество корма, потребляемого птицей за сутки и его энергетическая стоимость, т. е. так называемая большая энергия, GE. Определение суточного рациона — самая трудная задача во всей работе. До сих пор оно осуществлялось отстрелом птиц, особенно в тот момент, когда птица уходит на ночевку под снег. В этот момент птица имеет в зобу то количество корма, которого должно хватить до следующей кормежки. Наблюдения в природе показывают, что в большинстве случаев этот корм полностью расходуется ко времени выхода на утреннюю кормежку. Зная количество этого корма, достаточного для данного отрезка времени, и предполагая, что потребность в корме в течение суток более или менее равномерна (кишечник тетеревиных птиц работает зимой непрерывно, в особенности слепые кишки), можно экстраполировать эту потребность на все сутки (Лобачев, Щербаков, 1936; Семенов-Тян-Шанский, 1959). Количество корма в зобу при уходе на ночевку может быть определено только путем отстрела или отлова птицы. Энергия корма определяется его прямым калориметрированием. Недостатки этого метода: необходимость отстрела птиц, что нежелательно и не всегда возможно, а также необязательное расходование корма к утру, что в единичных случаях обнаружено мною у Tetrao urogallus.

Глухари - самец и самка.
2. Суточное количество экскрементов и их калорийность, дающие величину экскреторной энергии (ЕЕ). Суточное количество экскрементов определяется сбором экскрементов как тонкого, так и слепого отделов кишечника в подснежных камерах, на местах отдыха и кормежек. Напомним, что экскременты тетеревиных птиц выделяются: а) в виде твердых, цилиндрической формы «колбасок», представляющих собой главным образом спрессованную в прямой кишке клетчатку, прошедшую весь пищеварительный тракт и миновавшую входные отверстия слепых кишок, и б) в виде полужидкого вязкого вещества желтовато-коричневого цвета. Последнее выделяется слепыми кишками, как правило, раз в сутки, в момент оставления места ночлега. Время пребывания птиц в камерах определялось поначалу только прямыми наблюдениями. Впоследствии эти наблюдения, полученные в достаточном числе, позволили мне установить, что скорость выделения твердых экскрементов зимой примерно одна и та же у всех видов тетеревиных птиц, независимо от времени суток и температуры окружающей среды и составляет в среднем 11—12 мин на 1 экскремент. Это дало возможность, во-первых, по одной пробе экскрементов данной особи установить их суточную массу, учитывая, что в сутки в среднем выделяется 120 экскрементов. Проверка этого факта проводилась мной неоднократно, особенно в тех случаях, когда птица пребывала в одном месте более суток и удавалось собрать все суточное количество экскрементов. Указание на то, что в ночное время скорость выделения твердых экскрементов замедляется (West, 1972), моими данными пока не подтвердилось. При наблюдениях в природе следует учитывать, что экскременты слепых кишок, если они оказываются в подснежной камере или около нее, как правило, указывают на то, что птица покинула место ночевки утром, в спокойном состоянии. Отсутствие таких экскрементов говорит о том, что камера эта — либо место дневного отдыха, либо покинута птицей внезапно.
Калорийность экскрементов также определялась прямым калориметрированием. Накопившиеся данные позволяют установить, что калорийность основных зимних кормов тетеревиных птиц примерно одна и та же и варьирует в сравнительно небольших пределах (18.4—23.4 кДж/г). То же самое относится и к калорийности твердых экскрементов (20.1—21.3 кДж/г). В то же время калорийность экскрементов слепых кишок менее постоянна и зависит от вида птицы и степени суровости зимы.
Как было показано ранее, экскременты слепого кишечника содержат ряд хорошо усваиваемых веществ, прежде всего свободные жирные кислоты, но чем суровее условия зимы, тем более полно усваиваются они птицей, а следовательно, тем меньше становится и масса, и калорийность этих экскрементов.
Узнав общую энергию, поступившую в организм птицы, и экскреторную энергию, мы легко устанавливаем величину метаболизированной энергии (ME) по простой формуле:
ME = GE — ЕЕ. (1).
Отсюда уже по следующей формуле легко определить суточный бюджет энергии (DEB):
DEB = ME + mE1 (2)
где m — среднее уменьшение массы тела за сутки, а Е - калорический эквивалент 1 г этой массы.
Оценивая возможности этого метода, мы видим, что наиболее уязвимое его место — определение величины суточного рациона и, отсюда величины GE. Поэтому крайне важно разработать эффективные методы определения коэффициента усвоения пищи (МЕС). Значение последнего дает возможность определения GE только по экскреторной энергии по формуле (3):
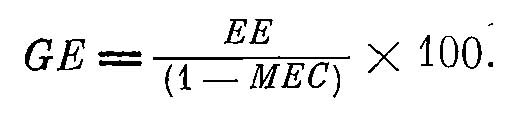
Формула 3.
Экскреторная же энергия, как мы видели, определяется наиболее точно.
Для определения МЕС наметились две возможности. Первая — приблизительное определение средних значений МЕС по прямым данным, полученным при содержании в неволе и методом отстрела. Кажущийся наиболее идеальным метод определения МЕС в условиях вольерного содержания, до сих пор невыполним по причине крайней трудности содержания тетеревиных птиц на естественных кормах. До сих пор это еще никому не удавалось без того, чтобы птицы не начали быстро истощаться (исключение — опыты с канадской дикушей; Pendergast, Boog, 1971а). Поэтому в рацион вольерных птиц сразу же начинают вводиться легко усваиваемые пищевые концентраты, что, конечно, делает невозможным определение коэффициента усвоения натурального корма. Еще одно исключение —остроумный эксперимент Мосса (Moss, Parkinson, 1972), где в опытах с Lagopus lagopus scoticus использовались передвижные вольеры. Экспериментальные птицы питались естественным кормом — вереском, находясь в вольерах, которые медленно передвигались по вересковому полю по мере потребления птицами этих растений. Среди остальных тетеревиных птиц эта методика может быть применена зимой только для полынного тетерева Centrocercus urophasianus, также питающегося зимой исключительно наземной травянистой растительностью (черная полынь).
Все полученные к настоящему времени данные показывают, что МЕС у различных видов тетеревиных птиц в зимнее время изменяется в пределах, от 29 до 45% (Pulliainen et al., 1968; Moss, 1972a; Андреев, 1980, и др.). Он может изменяться в течение зимы у одних и тех же птиц, но в среднем в середине зимы при температуре —10-—15 °С составляет около 31—33 %.
Второй путь — обнаружение такого маркера в пище, который, минуя пищеварительный тракт птицы, весь выводится с экскрементами. Зная его содержание в экскрементах и в корме, легко вычислить и количество последнего по одним лишь экскрементам по формуле (4):
DR = xэкс / xn формула (4)
где DR — сухая масса дневного рациона, xэкс — количество маркера в суточной порции экскрементов, а x
В настоящее время в качестве наиболее простого метода, не связанного с отстрелом птиц и дающего удовлетворительные результаты, можно предложить способ, основанный на определении экскреторной энергии и определении по ней метаболизированной энергии с помощью МЕС, вычисленного однажды экспериментально и достаточно постоянного у данного вида для данного вида корма и при данной температуре среды. Причем изменения величины МЕС, происходящие в течение зимы, можно определить по формуле:
MEC = Y – (EEс / GE) формула (5)
где Y — величина постоянная и в свою очередь вычисляется по формуле:
Y = 1 – (EEт / GE) формула (6).
В окончательном виде формула выглядит так:
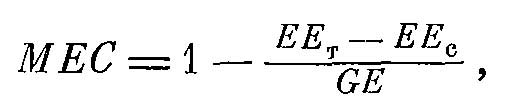
Формула 7.
где EEт — энергия твердых экскрементов, а EEс — энергия экскрементов слепых кишок.
Совершенно иной путь для вычисления суточного бюджета энергии (DEB) — расчет его по суточному бюджету времени (DTB) с помощью специальных коэффициентов, позволяющих выражать различные виды активности птицы в единицах базального метаболизма (Дольник, 1980). В таком случае расчет DEB выполняется по формуле:
DEB = (K1t1 + K2t2 + ... + Kntn) + htк(Tк — Tнк) + htа(Tа — Tт) + WпEп + FWр(Tт — Tа) формула (8)
где K1, K2 . . . — энергетические затраты на данный вид деятельности в единицах базального метаболизма (кДж/ч), t1, t2,… — временные затраты на данные виды деятельности, h — удельная теплоемкость (кДж/градус/час), Тк — температура подснежной камеры, Тнк — нижняя критическая температура птицы, tк — время пребывания в подснежной камере, Tа — температура наружного воздуха, Tт — температура тела птицы, tа — время пребывания вне снежной камеры, Wп — ежесуточная потеря массы тела, Eп — энергия этой потери (кДж/г), F — удельная теплоемкость корма, — масса суточного рациона в сыром виде. В этом расчете мы учитываем всю энергию, затрачиваемую птицей на жизнедеятельность, согревание корма до температуры тела и терморегуляцию. Отрицательный баланс массы показывает, что птицей тратится вся полученная из суточного рациона энергия, т. е. в этом расчете полностью представлен весь ежедневный энергетический бюджет птицы.
Единственным недостатком этого метода являются большие трудности определения величины базального метаболизма тетеревиной птицы эмпирически. Все полученные таким путем данные вызывают сомнения в своей достоверности по той причине, что для измерения ВМ необходима, как известно, птица в состоянии покоя и с пустым пищеварительным трактом. Однако известно, что в течение всей зимы слепые кишки тетеревиных птиц содержат большое количество пищевого экстракта и там непрерывно идет сложный пищеварительный процесс. Неудивительно поэтому, что экспериментальные попытки определения базального метаболизма у тетеревиных птиц зимой, предпринятые без учета этого обстоятельства, дали значение в 1.16 раза больше, чем дает теоретическая формула Кенди для неворобьиных птиц (West, 1972; Kendeigh et al., 1977). Освободить слепые кишки от химуса, не нарушая нормального физиологического состояния птицы, невозможно — для этого необходимо содержать птицу без корма по крайней мере несколько суток. Поэтому в данном случае следует, видимо, либо вычислять ВМ по формулам, выведенным для неворобьиных птиц (Kendeigh et. al., 1977), либо же использовать коэффициенты, выражаемые в единицах стандартного метаболизма. Во всяком случае в наших расчетах энергетические затраты птиц, находящихся в подснежных камерах ночью, оцениваются как и дневной покой, в 1.2 раза превышающие теоретически рассчитанную величину ВМ. Конечно, заманчиво получить прямые данные по величине обмена в подснежной камере на основании оставляемых там экскрементов, но это было бы возможно только в том случае: если бы слепой отдел кишечника работал абсолютно равномерно в течение суток. Однако противоречивые результаты, полученные на основании этих данных (Андреев, 1980), и сам факт значительных изменений степени наполненности слепых кишок в течение суток показывают, что никакой равномерности здесь нет.
В результате применения двух различных методик (по GE, ЕЕ, а также по DTB) были получены результаты, представленные в табл. 16. Для самцов Tetrao urogallus расчет суточного потребления энергии был выполнен и по определению зольного остатка в корме и в экскрементах. Результат примерно тот же — 1630.96 кДж/особь/сут (см. табл. 16). Прежде всего мы видим, что они дали близкие результаты, которые, видимо, достоверны. Во-вторых, реальная среднесуточная температура среды, в которой живет тетеревиная птица, заметно выше атмосферной. И, наконец, сравнение с величиной энергии существования, определенной для ряда фазановых птиц, показывает, что энергия существования куриных птиц независимо от их систематической принадлежности при равных массе и температуре среды одна и та же (Гаврилов, 1980). Это означает отсутствие у тетеревиных птиц особых энергетических адаптаций на тканевом уровне, и, следовательно, их успешное существование в условиях северной зимы, гибельных для фазановых, обеспечивается исключительно описанными выше адаптациями морфофизиологического и поведенческого плана.
Оценивая все адаптивные особенности тетеревиных птиц, связанные с зимним сезоном, мы видим, что эти птицы практически «сняли» для себя основные зимние проблемы — нехватку корма и мороз. Проблему корма решила способность к питанию так называемыми веточными кормами, имеющимися обычно в изобилии, а температурную проблему снимает способность сооружать термические убежища в снегу — явление само по себе уникальное в классе птиц.
Все адаптации тетеревиных птиц, связанные с зимним сезоном, являются весьма важными и взаимосвязанными, но по своей значимости они далеко неравнозначны. Основной, ведущей адаптацией было развитие способности усваивать грубый веточный корм, способности, которая в зачаточном состоянии существует и у фазановых птиц. Эта способность явилась результатом развития обширного слепого отдела кишечника, позволившего эффективно использовать питательные возможности грубой древесной пищи путем пролонгации процесса пищеварения в 10—60 раз. По сути дела слепой кишечник тетеревиных птиц — это своеобразный энергетический реактор, работающий непрерывно в течение всего зимнего сезона.
Оценивая роль снежного покрова в жизни тетеревиных птиц, мы должны признать его явно положительную, а не отрицательную роль. Именно снег дает им термические убежища — один из главных факторов, обеспечивающих положительный энергетический баланс птицы в течение зимы. Одновременно снег делает их менее уязвимым для хищников. Отрицательное воздействие снежного покрова усматривалось обычно в том, что он резко уменьшает кормовую базу птиц, скрывая наземный корм. Однако в предыдущих разделах было показано, что сезонная смена кормов, хоть и совпадает часто с установлением или исчезновением снежного покрова, им не определяется.
Для фазановых птиц в отличие от тетеревиных снег имеет только отрицательное значение. Он скрывает их основной корм, резко увеличивает потерю тепла через неоперенные лапы, попадает в незащищенные ноздри. Как правило, фазановые птицы не зарываются в снег даже при наличии такой возможности. В частности, это ни разу не было замечено у уларов. Но есть и исключения. Птицы из наиболее северных популяций Perdix perdix зарываются на ночь в снег совершенно так же, как и тетеревиные птицы (Westerskov, 1965; мои данные). Северные популяции уссурийского фазана также способны зарываться в рыхлый снег в случае опасности (Слепцов, Горчаковская, 1952), хотя способность их ночевать под снегом не доказана. Но даже при ночевке в подснежной камере фазановая птица должна получать значительно меньший энергетический выигрыш по сравнению с тетеревиной. Открытые, не защищенные оперением ноздри мало приспособлены для удержания влаги, что должно вызывать оледенение камеры, а отсутствие оперения плюсны сильно увеличивает затраты на терморегуляцию. Поэтому менее совершенная теплоизоляция фазановой птицы создает угрозу подтаивания снега в камере и намокания оперения. Эти начальные стадии освоения снежного покрова не могут поддерживаться отбором по причине свободного генетического обмена с остальными популяциями, зимующими в бесснежных условиях. Ни один из видов фазановых птиц не смог приспособиться полностью к снежным условиям, поскольку всегда имел возможность к отступлению в пределах ареала. Это заставляет предполагать, что для родоначальной формы тетеревиных птиц такое отступление было невозможно или по крайней мере резко ограничено.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
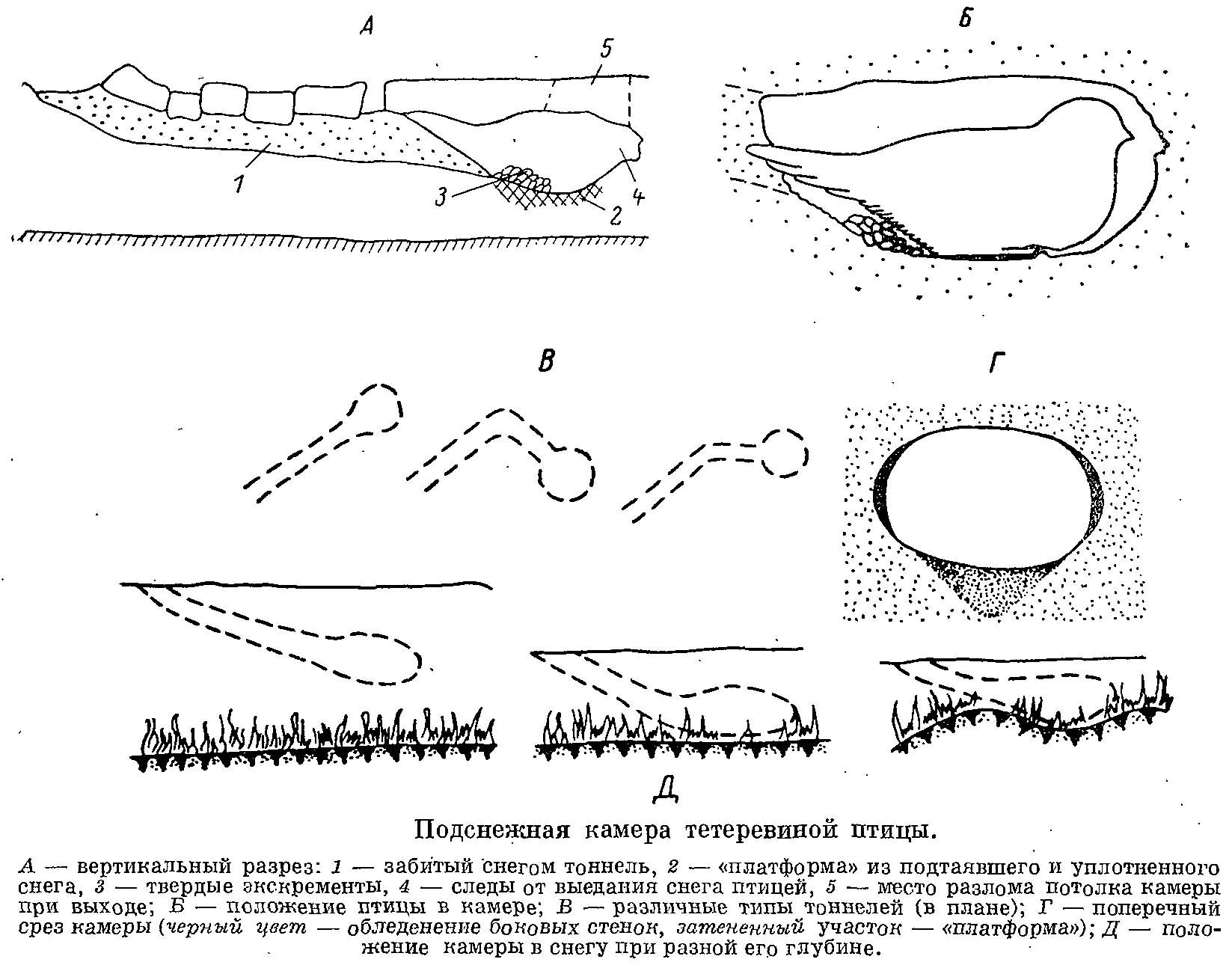 Использование снежного покрова тетеревиными птицами.
Использование снежного покрова тетеревиными птицами. Окунеобразные Чёрного моря. Отряд perciformes.
Окунеобразные Чёрного моря. Отряд perciformes.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
.") Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты).
Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты). Гнездование тетеревиных птиц.
Гнездование тетеревиных птиц. Брачный ритуал тетеревиных птиц - токование.
Брачный ритуал тетеревиных птиц - токование. Три дня с дрофой наедине.
Три дня с дрофой наедине. Птица стенолаз.
Птица стенолаз.
