Слепой отдел кишечника тетеревиных.
Тетеревиных птиц отличает от всех остальных представителей отряда большая длина кишечника и особенно сильное развитие его слепого отдела (Maumus, 1901; Schumacher, 1922, и др.). Первое детальное микроскопическое описание слепых кишок у 4 видов тетеревиных птиц дал Шумахер (Schumacher, 1922), но особенное внимание на них было обращено только в последние 10— 15 лет. Мною были просмотрены и макроскопически изучены слепые отделы кишечника всех видов тетеревиных птиц Палеарктики (за исключением Bonasa sewerzowi), а также В. umbellus, Falcipennis canadensis, Dendrogapus obscurus, Centrocercus urophasianus, Tympanuchus phasianellus и ряда видов сем. Phasianidae (Gallus gallus, Ammoperdix griseogularis, Alectorix kakelik, Tetraogallus himalayensis, T. altaicus, Perdix perdix, P. daurica, Coturnix coturnix, Perdicula asiatica, Francolinus pondicerianus, Phasianus colchicus).
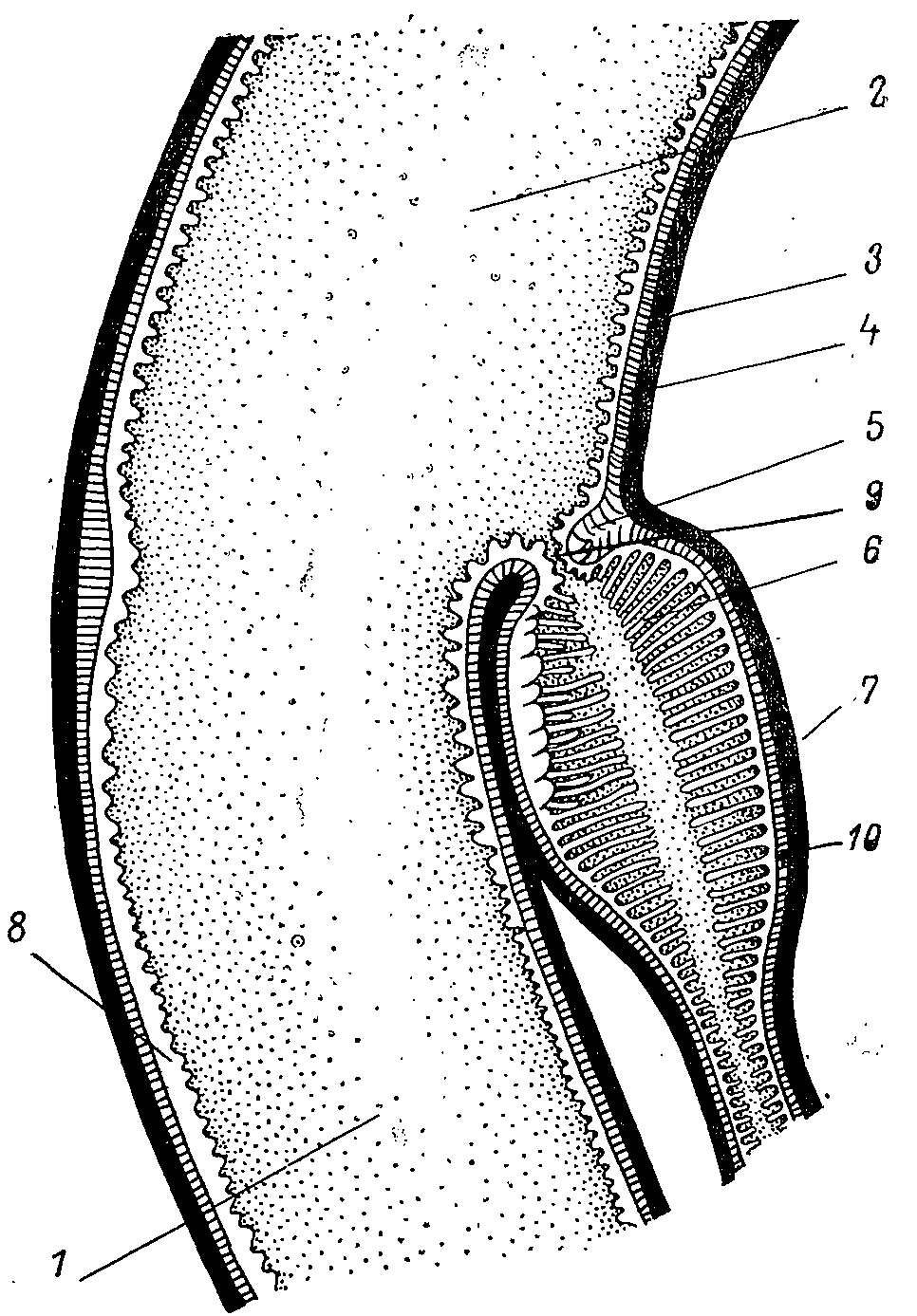
Рис. 1. Приемная капсула слепой кишки Tetrao urogallus. 1 — тонкая кишка; 2 — прямая кишка; 3 — наружный слой продольной мускулатуры; 4 — пестиковидные ворсинки прямой кишки; 5 — сфинктеры; 6 — ворсинки капсулы; 7 — приемная капсула; 8 — ворсинки тонкой кишки; 9 — фильтрующее устройство входа в приемную капсулу; 10 — внутренний слой поперечной мускулатуры.
Общее строение слепого отдела кишечника одинаково у всех видов тетеревиных птиц. Содержимое кишечника попадает сюда через приемную капсулу (рис. 1), вход в которую очень узок, замыкается сфинктером и имеет особый цедильный аппарат (Потапов, 1974). Вся внутренняя поверхность капсулы покрыта длинными булавовидными ворсинками. Часть дна капсулы и прилегающей наружной стенки занимает участок, покрытый крупными гребневидными выступами. Между ними и на некоторых гребнях располагаются те же булавовидные ворсинки, но на гребнях они очень короткие. Функция и микроскопическое строение этих образований пока неизвестны. Эти гребни занимают большие участки у Tetrao urogallus, Lyrurus tetrix, Bonasa bonasia и их очень мало у Lagopus mutus. Капсула обычно заполнена слизистым секретом, который полностью скрывает булавовидные ворсинки и оставляет просвет только в центральной части капсулы.
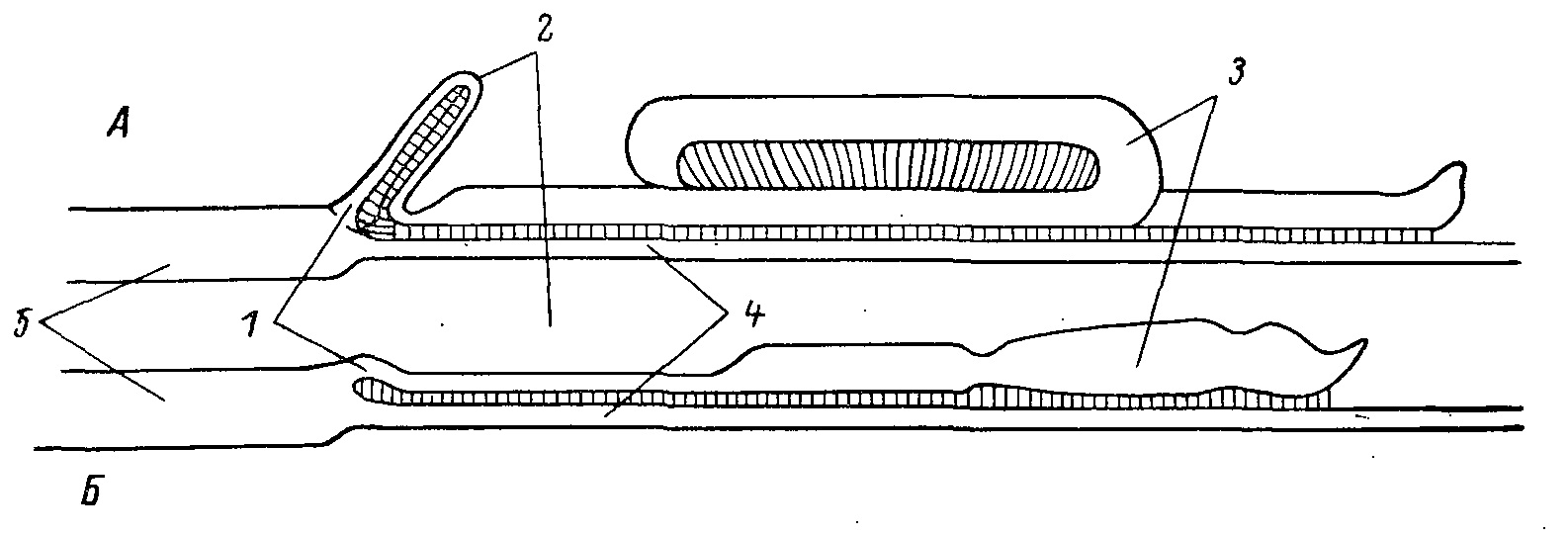
Рис. 2. Соединение слепых и тонких кишок. А — у тетеревиных; Б — у фазановых птиц. 1 — приемная капсула; 2 — «шейка» слепой кишки; 3 — «баллон»; 4 — тонкая кишка; 5 — прямая кишка.
Шейная часть отличается от остальных сильной мускулатурой с одинаковой толщиной как продольных, так и кольцевых слоев. Примечательно также развитие мерцательного эпителия, покрывающего как тонкие ворсинки, так и пространство между ними, но не сплошь, а отдельными участками (Schumacher, 1922).
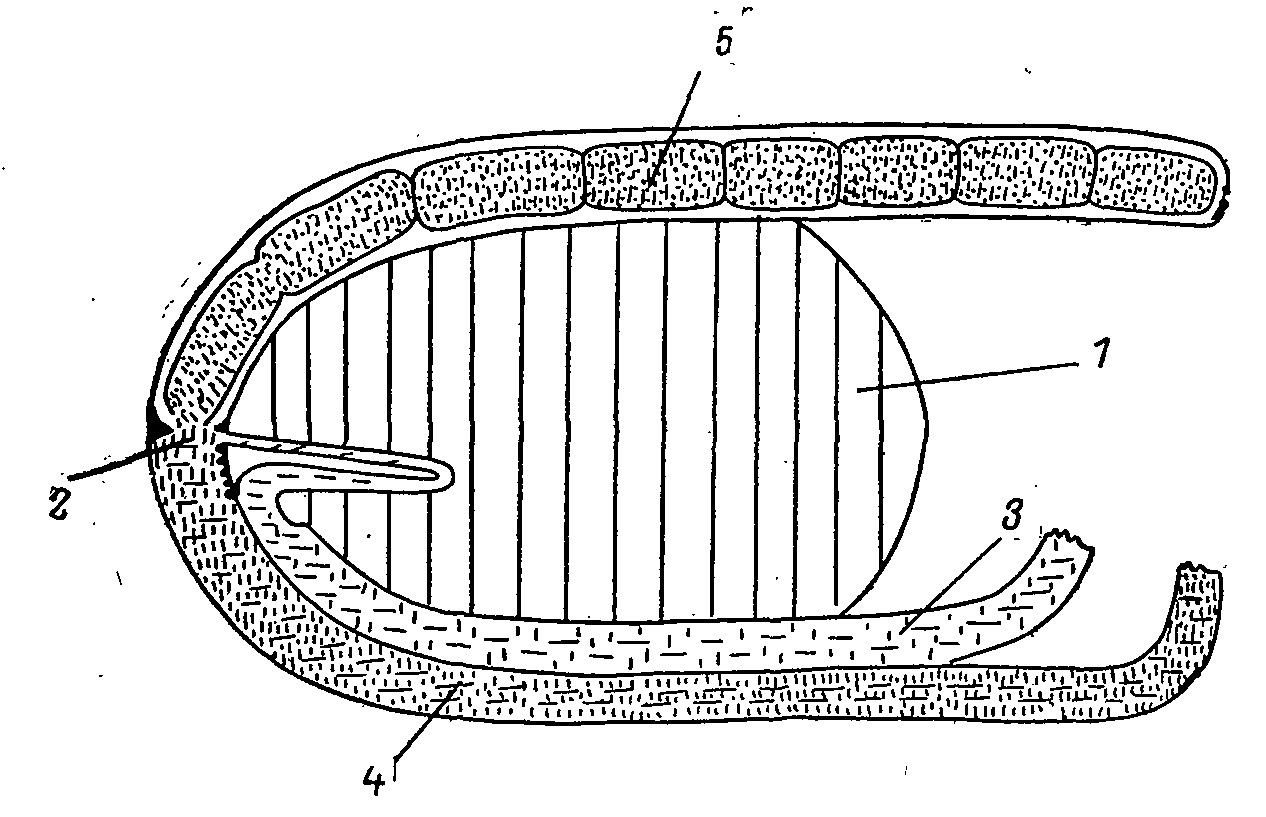
Рис. 3. Положение «распределителя» в полости тела (схема вертикального разреза). 1 — желудок; 2 — «распределитель»; 3 — слепая кишка; 4 — тонкая кишка; 5 — прямая кишка.
Главная часть слепой кишки — баллон в отличие от шейной части имеет мягкие и тонкие стенки. Характерная черта его строения — гребни из соединительнотканных образований, тянущиеся вдоль всей длины баллона и вдающиеся в его полость. Эти гребни, функция которых увеличение всасывающей поверхности, могут раздваиваться и число их на отдельных поперечных срезах колеблется от 7 до 10. Высота их у T. urogallus достигает 3—4 мм, у L. tetrix 2—3 мм, у L. lagopus и В. bonasia 1—2 мм. На микроскопических срезах хорошо видно, как от этих гребней в свою очередь отходят сильно ветвящиеся гребешки, покрытые всасывающим эпителием.
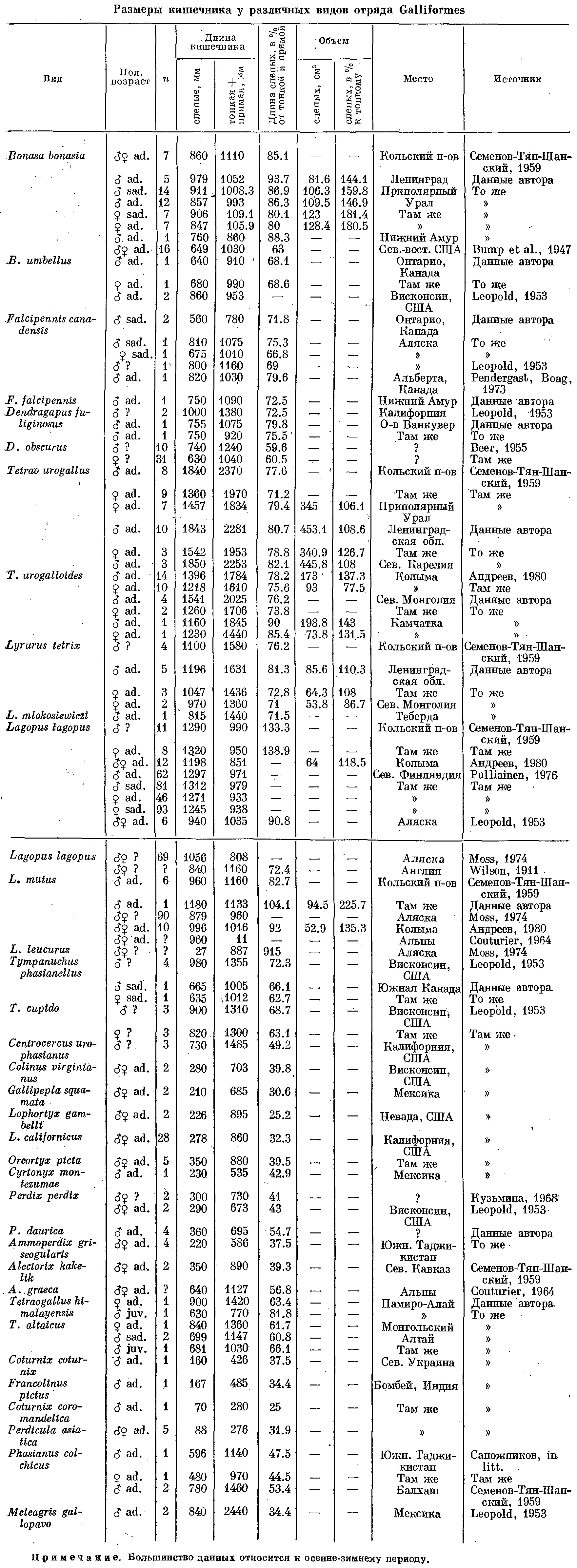
Таблица 1. Химический состав экскрементов слепого отдела кишечника Tetrao urogallus (27 I 1979 ), в %. Примечание: cостав свободных жирных кислот (100%): лауриновая 7.12, мипистиновая 17.13, пальмитиновая 24.43, олеиновая 51.32.
У различных видов Phasianidae, которые мне удалось осмотреть, слепой отдел кишечника устроен в основном по двум типам. Первый из них аналогичен тому, что мы видим у тетеревиных птиц, но гребни здесь развиты меньше и бывают хорошо выражены только в центральной части баллона. Кроме них есть еще короткие гребни, идущие в поперечном направлении и образующие с продольными как бы ячеистую сеть (Perdix, Coturnix; Fenna, Boag, 1974a, 1974b; мои данные). Другой тип строения встречен мною у уларов. В частности, у Tetraogallus altaicus продольных гребней нет вообще, а небольшие по высоте поперечные гребни образуют ячеистый рисунок.

Таблица 2. Площадь всасывающей поверхности тонких и слепых кишок у Tetrao urogallus (♂, Ленинградская обл., 1-я декада апреля).
В целом слепой отдел кишечника у фазановых и тетеревиных птиц не имеет принципиальных различий, но у первых он развит меньше, короток и прилегает по всей своей длине к тонкому кишечнику. У тетеревиных птиц резкое увеличение его длины вызвало образование двух петель. Одна из них — в шейной части, которая сложена вдвое, другая образована баллоном, средняя часть которого делает один виток (рис. 2).
Очень важным является положение «распределителя» — места, где соединяются слепой, тонкий и прямой отделы кишечника (Потапов, 1974). Он расположен на участке кишечника, ориентированного в вертикальной плоскости (рис. 3), а весь слепой отдел находится ниже уровня отверстий приемных капсул. Вследствие этого заполнение слепых кишок идет прежде всего под действием силы тяжести. Пищевая кашица, состоящая из смеси пищеварительного экстракта и твердых, непереваренных частиц, достигая распределителя, под влиянием перистальтики двигается дальше вверх, к прямой кишке. Жидкая фракция при этом заполняет любые встречные полости и проникает в любые отверстия. Таким образом она попадает сначала в складку тонкой кишки, к отверстиям приемных капсул, а затем и в сами приемные капсулы. Цедильный аппарат отверстий препятствует попаданию в капсулы крупных твердых частиц. Избыточное давление, возникающее в распределителе, повышается также действием клоакального сфинктера, задерживающего выход твердых экскрементов из прямой кишки наружу. Твердые частицы, попадающие в прямую кишку (это почти сплошь клетчатка, представленная сердцевиной побегов, перетертой хвоей, чешуйками сережек и почек и т. п.), здесь спрессовываются, отдают стенкам кишки почти, всю влагу и выводятся из организма в виде твердых экскрементов цилиндрической формы, резко отличаясь от вязких «жидких» экскрементов слепого отдела кишечника. Благодаря цедильному аппарату и малому размеру самих отверстий приемных капсул в слепые кишки попадает сравнительно гомогенная масса коричнево-желтого цвета, состоящая из пищеварительного экстракта и мельчайших частичек корма. Сфинктеры у приемных отверстий могут регулировать как поступление экстракта в слепые кишки, так и удаление из них содержимого. Как показали рентгеноскопические исследования, проведенные на Falcipennis canadensis, избыточному давлению в области распределителя способствует также противоположно направленная перистальтика тонкой и прямой кишок (Fenna, Boag 1974b).

Таблица 3. Размеры кишечника у различных видов отряда Galliformes. Примечание: большинство данных относится к осенне-зимнему периоду.
Функция мощной мускулатуры шейки слепой кишки, а также сфинктера распределителя не вполне ясна и, вероятно, связана с экскрецией. Несомненно, что для удаления содержимого слепых кишок прямой отдел кишечника должен быть пуст. Как показывают наблюдения, экскреция из слепых кишок происходит раз в сутки, рано утром, при оставлении птицей места ночевки. Обычно к этому времени весь запас корма, запасаемый с вечера, расходуется и кишечник за исключением его слепого отдела бывает пустым. Но в ряде случаев, как показали мои наблюдения, птица к утру расходует не весь запас корма: после оставления места ночлега и удаления экскрементов из слепых кишок, она вновь начинает равномерное выделение твердых экскрементов, еще не приступая к кормежке. Перед экскрецией из слепых кишок поступление содержимого тонкого кишечника в прямую кишку прекращается, что может быть осуществлено только сфинктером распределителя (рис. 1). Содержимое слепых кишок удаляется раз в сутки (в экстремальных ситуациях даже через день) за 3—4, иногда 5 сокращений кишечной мускулатуры таким же количеством порций. Время их выделения — утро — не случайно. Во-первых, вязкий и пахучий «выброс» (так мы будем называть для краткости эти экскременты) может испачкать оперение птицы, сидящей в снежной камере, а также демаскировать место ночлега. Во-вторых, перед утренней кормежкой необходимо освободить место для новой порции пищеварительного экстракта. В условиях неволи некоторые птицы выделяют экскреты из слепых кишок дважды в сутки, утром и вечером. Кроме того, выделение экскрементов из слепых кишок может иметь место и днем, но никогда — ночью.
Все, что известно на данный момент о слепых кишках тетеревиных птиц, явно указывает на то, что зимой они берут на себя основную часть пищеварительного процесса. До сих пор высказывается мнение, что основное значение слепых кишок — разложение клетчатки с помощью специальных желез и бактериальной флоры (Кузьмина, 1968; МсВее, West, 1969), хотя специальные исследования (Suomalainen, Arhimo, 1945) доказали, что незначительная бактериальная флора, способная разлагать клетчатку, имеется здесь примерно в таком же количестве, что и в тонком кишечнике. Характер содержимого всего кишечника показывает, что как раз почти вся клетчатка идет в прямую кишку, минуя слепой отдел. Согласно биохимическим анализам, твердые экскременты тонкого кишечника состоят на 85—95% из клетчатки. В экспериментах с Lagopus lagopus максимально усваивалось не более 27% клетчатки. Поэтому можно думать, что в основе очень сложного пищеварительного процесса, происходящего в слепых кишках тетеревиных птиц, лежат совсем иные процессы (Pulliainen et al., 1968). Бурная секреция эпителия слепых кишок, установленная Шумахером и подтвержденная изучением происходящей здесь ферментативной деятельности (МсВее, West, 1969), указывает на большую сложность этих процессов, в ходе которых происходит разложение и всасывание компонентов, предварительно экстрагированных при прохождении корма по пищеварительному тракту. Само прохождение корма по пищеварительному тракту у тетеревиных птиц длится при +4 °С около 4 ч (Bump et al., 1947). Перевод же пищеварительного экстракта в слепой отдел кишечника позволяет удлинить процесс пищеварения в несколько раз, на что первым обратил внимание Семенов-Тян-Шанский (1959). Никаких аналогов этому у других птиц неизвестно. Можно предполагать, что здесь происходит расщепление белков и жиров и всасывание продуктов расщепления, а также нейтрализация различных токсичных компонентов древесных смол.
Большой интерес представляет химический состав экскрементов слепых кишок. Предварительный самый общий анализ дал следующие результаты (табл. 6). Здесь привлекает внимание очень большой процент (до половины всего количества) липидного компонента. Более подробный анализ липидов экскрементов слепого кишечника Tetrao urogallus, выполненный методом тонкослойной хроматографии и при помощи хроматографа, показал очень высокий процент фосфолипидов и в меньшем количестве моноглицеридов. Примечательно присутствие, хоть и в небольшом количестве, ряда свободных жирных кислот, обычно хорошо усваиваемых организмом. Это подтверждает предположение, что слепой кишечник работает с большим запасом. В благоприятных условиях усваиваются далеко не все питательные вещества и часть выделяется вместе с отработанной массой перед очередным наполнением новой порцией экстракта. Чем суровее условия, чем ниже температура, чем дольше птица находится под снегом, тем полнее усваиваются питательные вещества в слепых кишках и тем меньше экскрементов они выделяют. У Т. urogallus, например, в зависимости от условий количество этих экскрементов изменяется в 6-кратном размере. В экстремальных ситуациях экскременты из слепых кишок вообще могут выделяться не каждый день (Bonasa bonasia; Андреев, 1980).
Таким образом, слепой отдел кишечника тетеревиных птиц — важное приспособление, позволяющее им с достаточной полнотой извлекать питательные вещества из грубого древесного корма. Это достигается резким увеличением (в несколько раз) всасывающей поверхности эпителия и значительным удлинением процесса переработки пищевого экстракта. В слепые кишки попадает экстракт, миновавший уже тонкий кишечник, и, не будь их вообще, он просто бы удалялся из организма, не успев отдать свои питательные вещества за сравнительно короткое время прохождения по кишечному тракту. Задержка химуса в слепых кишках увеличивает время переваривания каждой порции по крайней мере в 8 раз, а в ряде случаев, вероятно, и вдвое больше. Непрерывное круглосуточное функционирование слепых кишок в течение всей зимы постоянно снабжает организм необходимыми веществами, что позволяет тетеревиным птицам в норме обходиться без запасания дополнительных энергетических резервов в виде жировых отложений. Нельзя сказать, что у тетеревиных птиц их вообще нет, они бывают и иногда довольно значительные. Так, в ультраконтинентальных районах Верхоянья Tetrao urogalloides и Bonasa bonasia накапливают мощный слой жира в подкожных депо и на внутренностях (Егоров и др., 1959). Столь же жирны Lagopus mutus hyperboreus зимой на Шпицбергене (Johnsen, 1941). Заметные жировые накопления имеются поздней осенью у обыкновенного глухаря, обитающего в северной части таежной зоны, где низкие температуры устанавливаются раньше, чем снежный покров приобретает мощность, достаточную для зарывания птиц. Невозможность зарывания в снег резко повышает затраты птиц на терморегуляцию, и накопление к этому времени жирового резерва у тетеревиных птиц как по всему северу тайги вообще, так и в малоснежной Якутии в частности оказывается крайне необходимым. В большинстве же случаев работа слепых кишок вкупе с деятельностью тонкого кишечника полностью обеспечивают птиц необходимой энергией в зимнее время.
До сих пор еще не определено, какая доля из всей усвоенной энергии поступает из слепого отдела кишечника. Для этого необходимо точно знать, какое именно количество химуса вошло в слепую кишку и какая часть его была усвоена. Выяснить это в настоящее время вряд ли возможно, поскольку здесь происходит непрерывное перемешивание поступающих свежих порций с уже частично переваренным содержимым, а также с секретом, постоянно выделяемым клетками эпителия. Поэтому возможны только косвенные расчеты. В частности, оценка процесса ферментации в слепых кишках показывает, что здесь может усваиваться от 6 до 30 % энергии, поступающей с кормом, т. е. они дают 3—18% энергии существования (МсВее, West, 1969). Расчеты Андреева (1980) по разности массы экскрементов, выделяемых слепыми кишками зимой и весной, а также по скорости наполнения слепых кишок в течение суток дали близкие цифры порядка 30—42 % энергии существования. Мною выполнен косвенный расчет этой доли по соотношению всасывающей поверхности тонкого и слепого отделов кишечника. В этом расчете необходимо учитывать, что тонкий отдел кишечника, как и слепой, в течение зимы никогда не бывает пустым, но его наполненность к утру, да и в первые часы кормежки намного уступает таковой слепых кишок. Кроме того, каждая единица всасывающей поверхности тонкого отдела кишечника по сравнению со слепыми кишками должна работать менее эффективно, поскольку ворсинки здесь далеко не достигают таких размеров и сложности строения. Однако в расчете условно принимается, что каждая единица всасывающей поверхности в слепых и тонких кишках работает с одинаковой эффективностью, т. е. скорее принижается роль слепых кишок, чем завышается. Но даже при таких допущениях данные прямых измерений всасывающей поверхности различных отделов кишечника, проводившихся на свежедобытых самцах Tetrao urogallus, находившихся на зимней диете (сосновая хвоя), показывают, что на долю слепого отдела кишечника приходится почти 2/3 (65 %) от всей всасывающей поверхности тонкой и слепых кишок (табл. 2). Отсюда мы можем считать, что не менее 2/3 всей энергии, усваиваемой из пищи, дает птице именно слепой отдел кишечника.
В теплый сезон года слепые кишки уменьшаются в размерах, наполненность их минимальна, но все же пищеварительные процессы там идут. Регулярное употребление летом всеми тетеревиными древесных кормов, хоть и в малом количестве, явно поддерживает функционирование органа, на какой-то период становящегося ненужным. В зоне тундры у белых куропаток слепые кишки активно функционируют все лето (А. В. Андреев, устное сообщ.). Недавно была обнаружена еще одна исключительно важная функция слепых кишок. Было установлено, что мочевая кислота благодаря обратной перистальтике прямой кишки попадает в слепые, где имеется бактериальная флора, активно ее разлагающая и возвращающая азот в организм птицы (Mortensen, Tindall, 1977, 1981). Таким образом, стало понятным казавшееся неразрешимым противоречие между бедностью основных зимних кормов белковыми компонентами и хорошим состоянием птиц, существующих на этой диете.
Данные, приведенные в табл. 3, хорошо показывают степень развития слепых кишок у представителей тетеревиных и фазановых птиц. У фазановых длина слепых кишок значительно меньшая. По сути дела только улары, обитающие в наиболее суровых для фазановых птиц условиях, приближаются к нижнему пределу вариаций, наблюдаемых у тетеревиных. Хотя длина слепых кишок, например, у алтайского улара равна таковой L. lagopus scoticus, но последняя в 4 раза меньше его по массе тела, и если мы выразим длину слепых кишок в относительных цифрах, приходящихся на 1 кг массы птицы, то разрыв между представителями двух семейств будет вполне отчетлив.
Относительный размер слепых кишок, а также абсолютные размеры всего кишечника зависят от длительности и степени суровости зимнего периода, а также от длины ночного периода в середине зимы. Это наблюдается как у тетеревиных, так и у фазановых птиц. В настоящее время можно считать установленным, что хотя длина слепых кишок различна у разных видов, но в пределах вида она не является постоянной величиной и зависит от возраста птицы, от характера кормов, от сезона года и степени его суровости (Lewin, 1963; Pendergast, 1969; Moss, 1972; Pendergast, Boag, 1973; Fenna, Boag, 1974b).
Экспериментально было установлено, что длина слепого отдела кишечника Falcipennis canadensis зависит от состава пищи и чем больше в диете грубых, плохо усваиваемых кормов с преобладанием клетчатки, тем длиннее будут слепые кишки. В то же время у Coturnix coturnix длина всего кишечника и слепого отдела в особенности увеличивалась в эксперименте при пониженных температурах, но не зависела от состава корма (Fenna, Boag, 1974а).
Тот факт, что длина слепых кишок признак в известной степени сиюминутный и сугубо адаптивный, не мешает, однако, использовать его для характеристики сем. Tetraonidae. Сравнительно большая длина caeca определенно указывает на специфику питания в условиях низких температур. Возможно, что процесс увеличения слепого отдела у тетеревиных птиц может быть отчасти обратим’ но только отчасти, что хорошо доказывается экологической обусловленностью южных границ распространения семейства.
Функциональное значение caeca у фазановых птиц исследовано недостаточно, но при удалении слепых кишок у домашних кур и индеек в раннем возрасте птицы благополучно развивались дальше (Farner, 1960). Это явно говорит о крайне небольшом значении слепых кишок в жизни этих птиц. У взрослых кур с удалением caeca резко падала и без того небольшая способность к усвоению целлюлозы (Radeff, 1928). Кроме того, у кур здесь был обнаружен синтез витаминов (Couch et al., 1950). У видов, зимующих в более суровых условиях, caeca несомненно играет большую роль. Во всяком случае у кеклика из таджикской популяции, содержавшегося в домашних условиях, происходило регулярное выделение экскрементов слепых кишок утром и вечером (P. JI. Беме, устное сообщ.).
Таким образом, специализация на питании зимой грубыми древесными кормами вызвала у тетеревиных птиц целый ряд адаптивных изменений в органах пищеварения. Наиболее сильно они проявились в кишечнике, где возникла совершенно особая система, взявшая на себя основную роль в процессе пищеварения на время зимнего сезона. Эта система слепых кишок, в известной мере дублирующая работу тонкого кишечника, хорошо развита у всех куриных птиц, но в данном случае она взяла на себя совершенно новую функцию. Работа этого отдела у тетеревиных птиц обеспечивает непрерывное круглосуточное поступление энергии в организм, резко увеличивает эффективность пищеварительного процесса и обеспечивает к тому же кругооборот азота. Именно функционирование слепого отдела, позволяет тетеревиным птицам эффективно использовать грубые, бедные белками древесные корма, главным достоинством которых является их изобилие и доступность в течение всего зимнего сезона.
С морфологической точки зрения все различия пищеварительной системы тетеревиных и фазановых птиц имеют скорее количественный, чем качественный характер. Мы видим здесь только дальнейшее развитие тех или иных структур, которые имелись уже у фазановых птиц — предков тетеревиных. Ничего принципиально нового здесь не возникло. Единственное новшество — специфичный процесс пищеварения в слепых кишках — физиологического порядка, но возможно, что в дальнейшем нечто подобное будет обнаружено и у некоторых фазановых птиц, наиболее близких тетеревиным по образу жизни.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
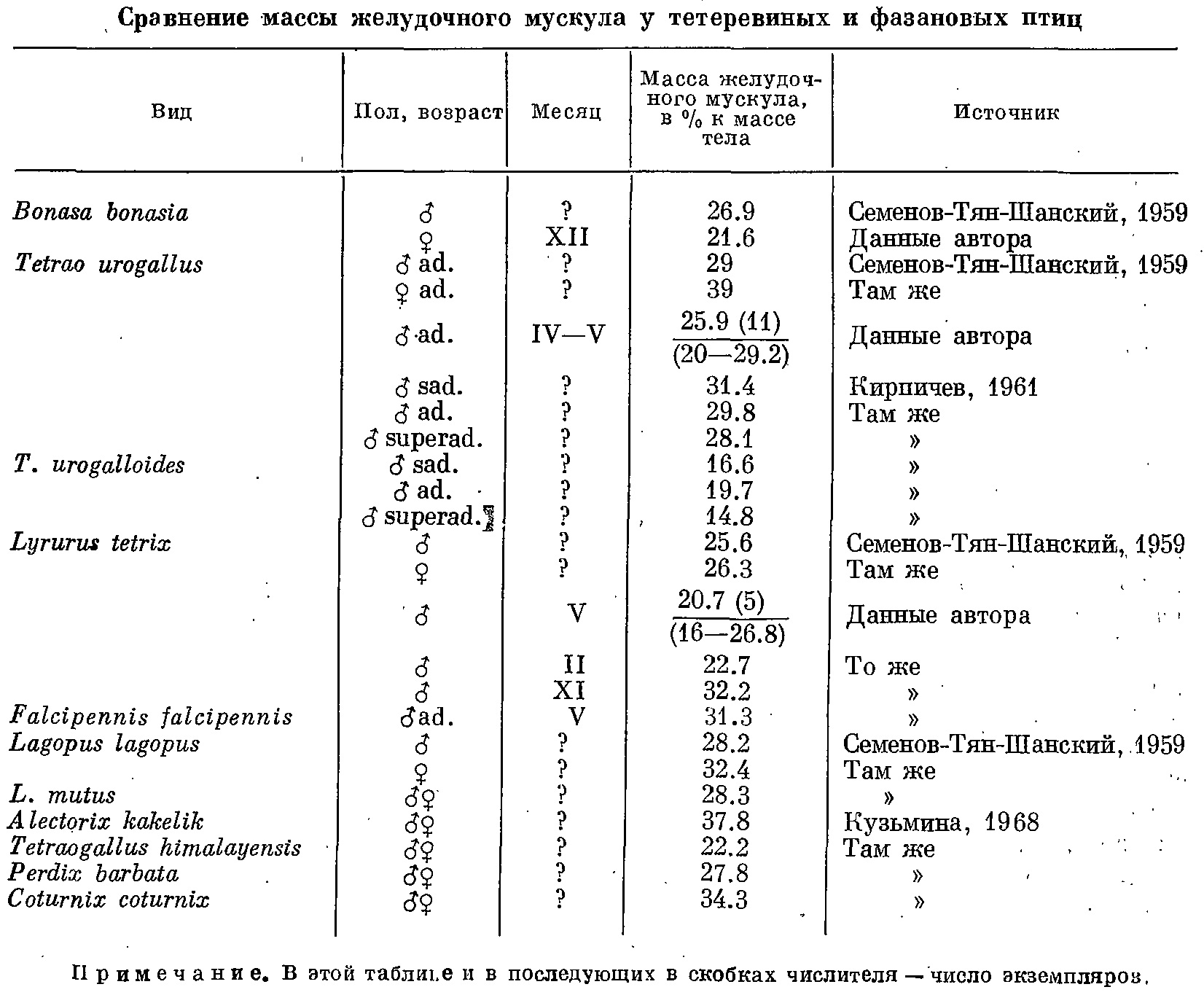 Мускульный желудок тетеревиных птиц.
Мускульный желудок тетеревиных птиц. Гибриды на воле.
Гибриды на воле.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Зоб тетеревиных птиц. Сколько еды может вместить зоб?
Зоб тетеревиных птиц. Сколько еды может вместить зоб? Рамфотека тетеревиных птиц. Как птицы пережёвывают растения.
Рамфотека тетеревиных птиц. Как птицы пережёвывают растения. Пищеварительная система тетеревиных птиц.
Пищеварительная система тетеревиных птиц. Скелет и мускулатура тетеревиных птиц.
Скелет и мускулатура тетеревиных птиц. Как кормить птенца и ухаживать за маленькой дикой птицей.
Как кормить птенца и ухаживать за маленькой дикой птицей.
