Скелет и мускулатура тетеревиных птиц.
В целом как скелет, так и мускулатура сем. Tetraonidae имеют сравнительно немного особенностей, но некоторые из них весьма существенны и целиком обусловлены спецификой семейства.
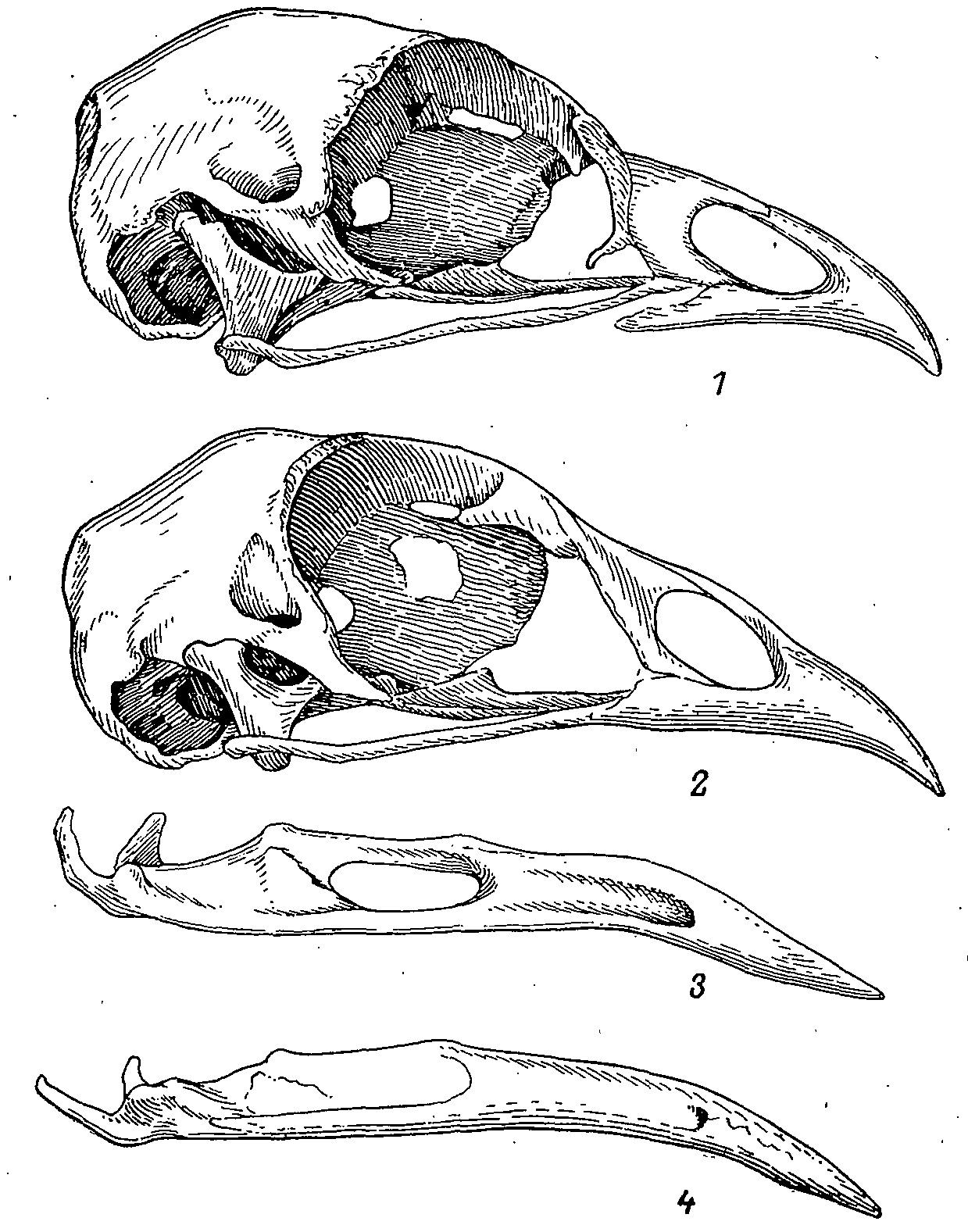
Рис. 1. Черепа тетеревиных (Lyrurus tetrix, 1, 3) и фазановых птиц (Phasianus colehicus, 2, 4). 1,2 — череп; 3, 4 — нижняя челюсть.
Общая конструкция черепа и его пропорции у тетеревиных птиц сходны с таковыми у фазановых. Основные отличия заключаются в следующем.
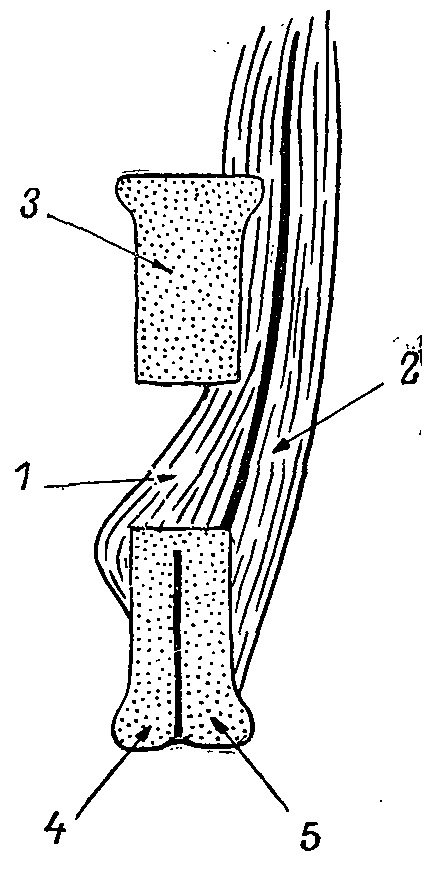
Рис. 2. Схема поперечного среза нижней челюсти Tetrao urogallus на уровне каудальной части foramen mandibularis. 1 — m. adductor mandibulae externa medialis; 2 — m. mandibulae externa superficialis; 3 — os supraangulare; 4 — os angulare; 5 — os dentale.
1. У тетеревиных птиц резко дифференцирована прокинетическая гибкая зона, которая ориентирована относительно конька надклювья значительно круче, чем у фазановых, особенно ее латеральные участки. Такая ориентация позволяет сдвинуть рабочий участок клюва ближе к его основанию, что дает определенный выигрыш в силе (Гамбарян, 1978) (рис. 1). Значение такого выигрыша, учитывая специфику зимнего питания тетеревиных птиц, вполне очевидно.
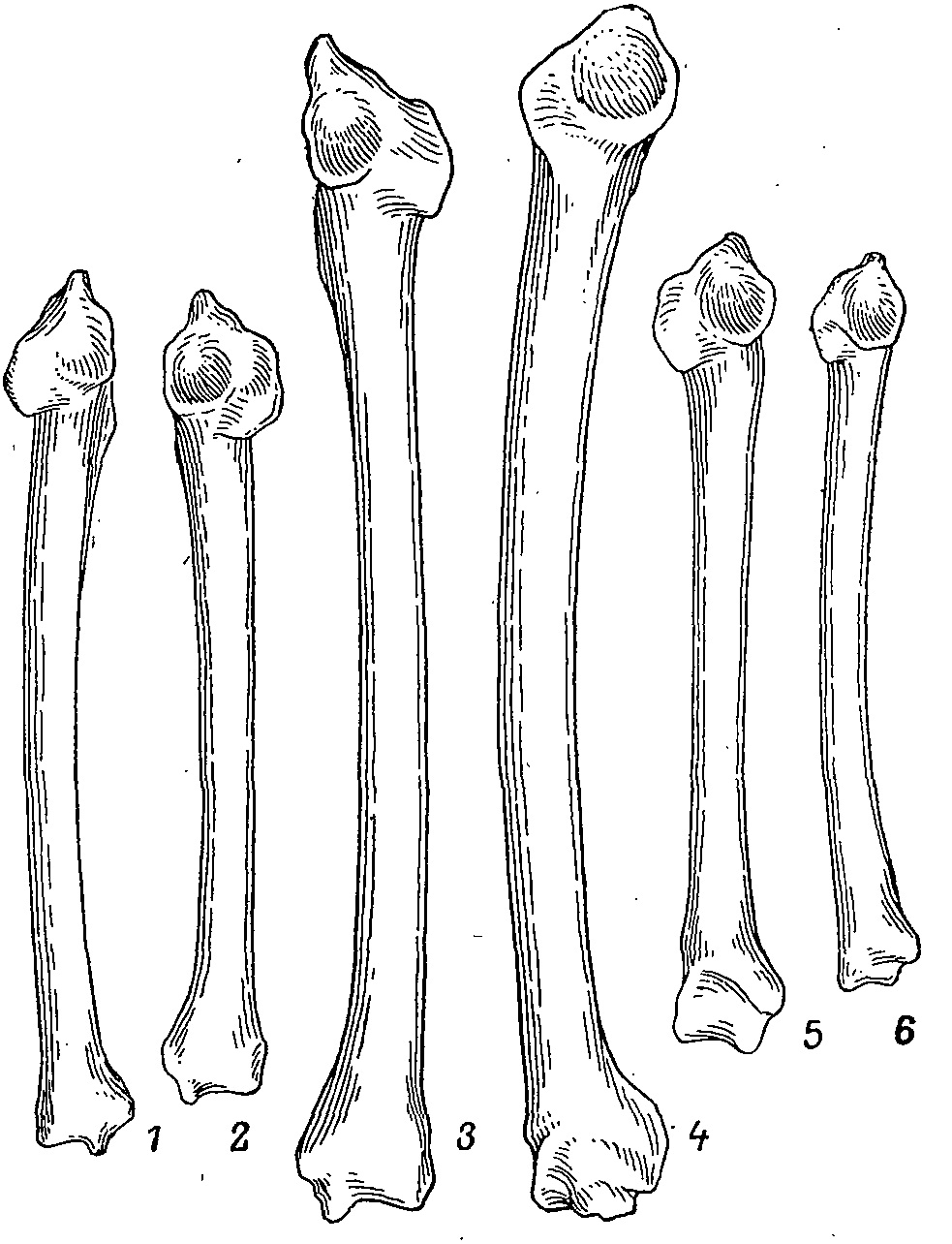
Рис. 3. Плечевые кости куриных птиц. 1 — Lagopus lagopus, 2 — Tympanuchus phasianellus, 3 — Tetrao urogallus; 4 — Crax spp.; 5 — Tetraogallus himalayensis; 6 — Phasianus colchicus.
2. Усилена мускулатура, приводящая нижнюю челюсть и опускающая надклювье, хотя принципиальных отличий от фазановых в расположении и взаимоотношениях челюстных мускулов нет — просто у тетеревиных птиц находит свое крайнее выражение работа челюстей по принципу ножниц, где помимо вертикальных движений они совершают еще встречные движения в горизонтальной плоскости. Эффективность таких движений обеспечивается более ростральным креплением наружного аддуктора (м. adductor mandibulae externus) на осевом черепе (Дзержинский, 1977). Эти особенности тоже связаны с характером зимнего корма, требующего гораздо больших усилий на отделение его от ветвей, чем при питании нежными частями растений.
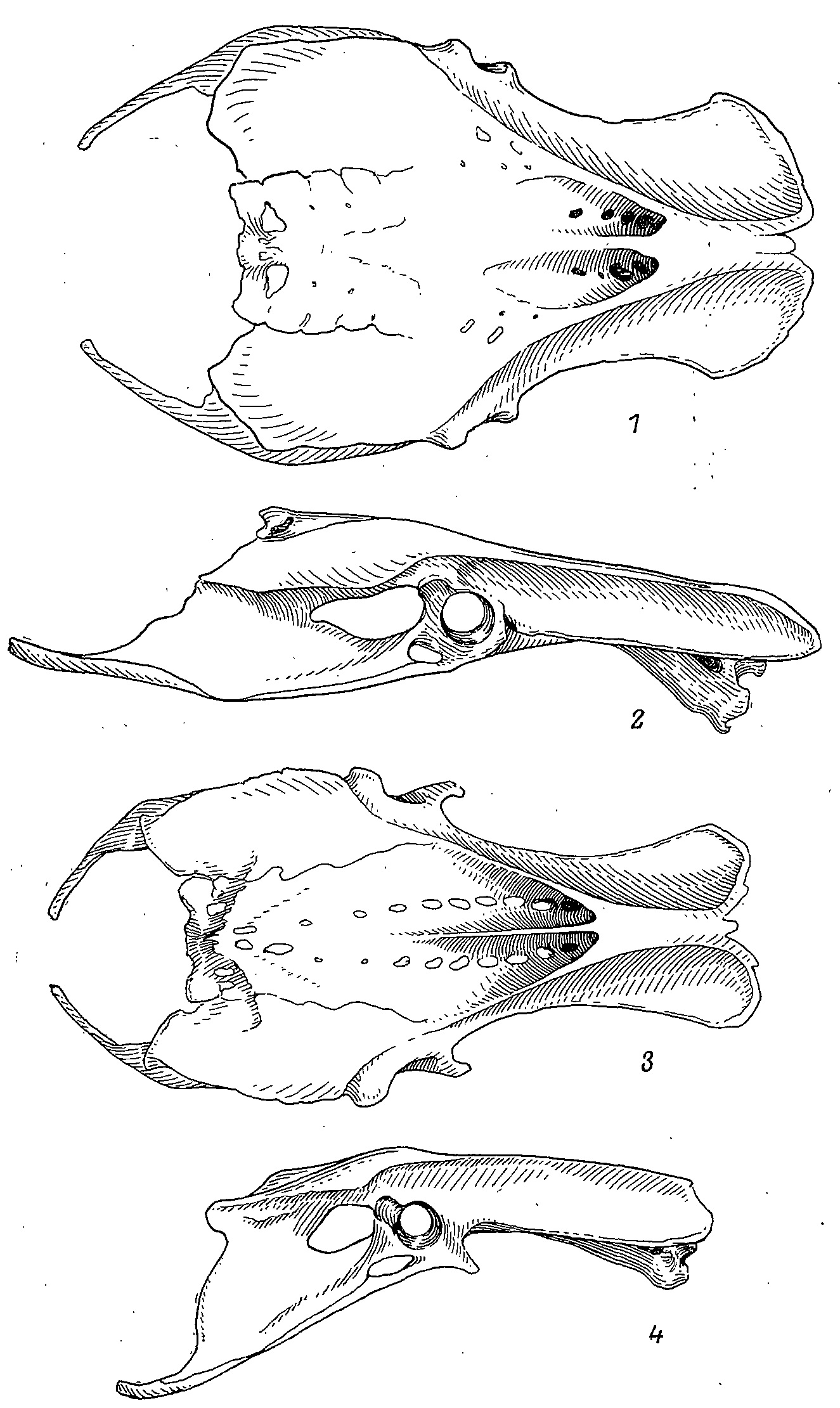
Рис. 4. Таз тетеревиной (Tetrao urogalloides, ♀, 1, 2) и фазановой (Phasianus colchicus, ♂, 3, 4) птиц. 1,3 — вид сверху; 2, 4 — вид сбоку.
3. С усилением челюстной мускулатуры связано значительное развитие нижнечелюстного отверстия, f. mandibulare, о чем коротко упоминает Фюрбрингер (Furbringer, 1888), но в последних морфо-функциональных исследованиях этот факт даже не упоминался. Между тем наличие этого отверстия явно связано с усилением m. a. mandibulae externus и необходимостью более прочного его крепления на нижней челюсти (рис. 2). У большинства фазановых птиц это отверстие отсутствует, хотя у некоторых видов (Tetraogallus caucasicus, Gallus ssp.) кость здесь сильно утоньшена и окружена небольшими гребнями. F. mandibulare таких же размеров, что и у тетеревиных птиц, я обнаружил у представителей родов Perdix, Coturnix, Rollulus, где функциональная обусловленность его не ясна.
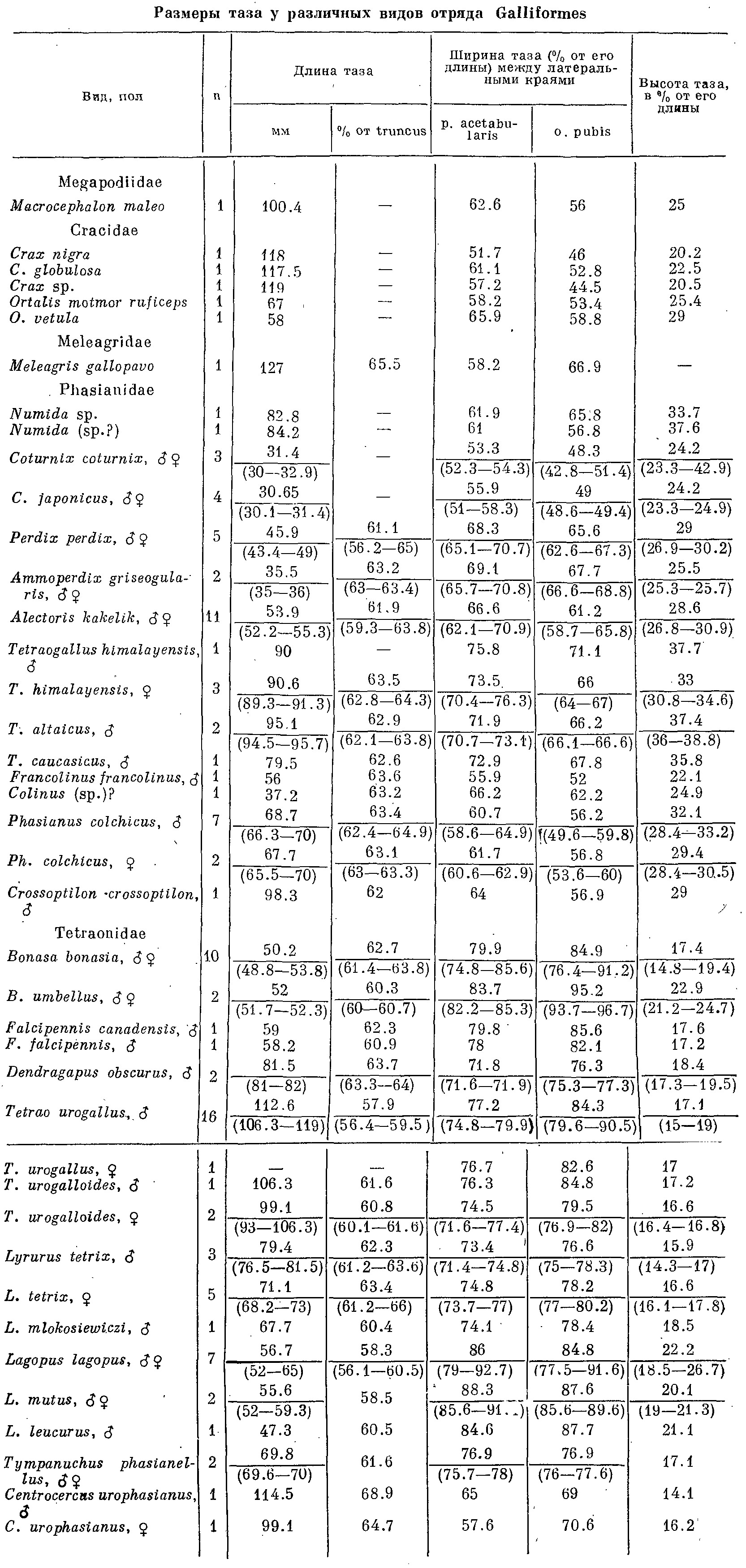
Таблица 1. Размеры таза у различных видов отряда Galliformes. Примечание: в этой таблице в числителе — средние величины, в знаменателе даются пределы колебаний.
В целом можно заключить, что в пределах сем. Tetraonidae строение черепа значительно более однообразно, чем в сем. Phasianidae. Среди первых единственным отклонением являются виды рода Tetrao, где в строении черепа имеется ряд глубоко специализированных черт. Это относится прежде всего к самцам, они прослеживаются вполне отчетливо. Грубый зимний корм и необходимость употребления его в больших количествах (из-за крупных размеров птиц) обусловили уникальную структуру глухариного черепа — вытянутого, клинообразного в плане и одновременно уплощенного в дорсовентральном направлении, с мощной крышей и рядом других особенностей. Развитие такой массивной конструкции в онтогенезе идет довольно медленно, оканчиваясь у самцов Т. urogallus не ранее, чем к 4-му году. Долгий рост костей как осевого, так и висцерального отделов черепа препятствует их раннему сращиванию и то, что у остальных тетеревиных птиц проходит на ранних стадиях онтогенеза, здесь задерживается до более позднего возраста. Замедление срастания костей, «всплывание» в онтогенезе некоторых ранних стадий, уже выпавших из развития других видов (например, стадия, на которой различим даже меккелев хрящ; Кириков, 1939), могут рассматриваться как пример неотенического сброса специализации и развития на исходной основе новой своеобразной конструкции. Именно по этой причине нельзя рассматривать череп глухарей как типичный для семейства. Наоборот, он как раз наименее типичен.
Пояс передних конечностей не имеет сколько-нибудь специфичных черт, свойственных исключительно этому семейству, хотя некоторые особенности и получают у них преимущественное развитие. Так, например, у всех тетеревиных птиц имеется хорошо выраженное пневматическое отверстие на лопатке (на мостике между акроминоном и гленоидальной фасеткой). У фазановых птиц этого отверстия нет, за исключением Pavo cristatus (Holman, 1964). В качестве другого примера можно привести локтевой выступ плечевой кости, который у тетеревиных птиц заметно выдается за ее нижнюю поверхность (рис. 3). Однако такую же ситуацию мы наблюдаем у некоторых краксов (Ortalis). Пропорции же различных элементов плечевого пояса подвержены значительной изменчивости даже в пределах одного вида (длина плеча у самок Perdiz daurica в пределах 59—66.5 % от truncus; Кузьмина, 1977). В целом пропорциональные соотношения элементов плечевого пояса всецело определяются образом жизни вида и не отражают родственных связей последних (Штегман, 1950). Характеризуя морфологию кисти тетеревиных птиц, Штегман (1979) отметил отсутствие когтя (второй фаланги) первого пальца, наличие развитой связки lig. humero-carpalis и слабое развитие m. flex. sublimis. Но и по этим признакам отличия фазановых и тетеревиных птиц нечеткие, поскольку коготь первого пальца мал у Perdix, Alectoris, Tetraogallus и полностью отсутствует у Crossoptilon. У этих родов найдены начальные стадии развития lig. humero-carpalis, отсутствующей у Gallus, Phasianus и Meleagris.
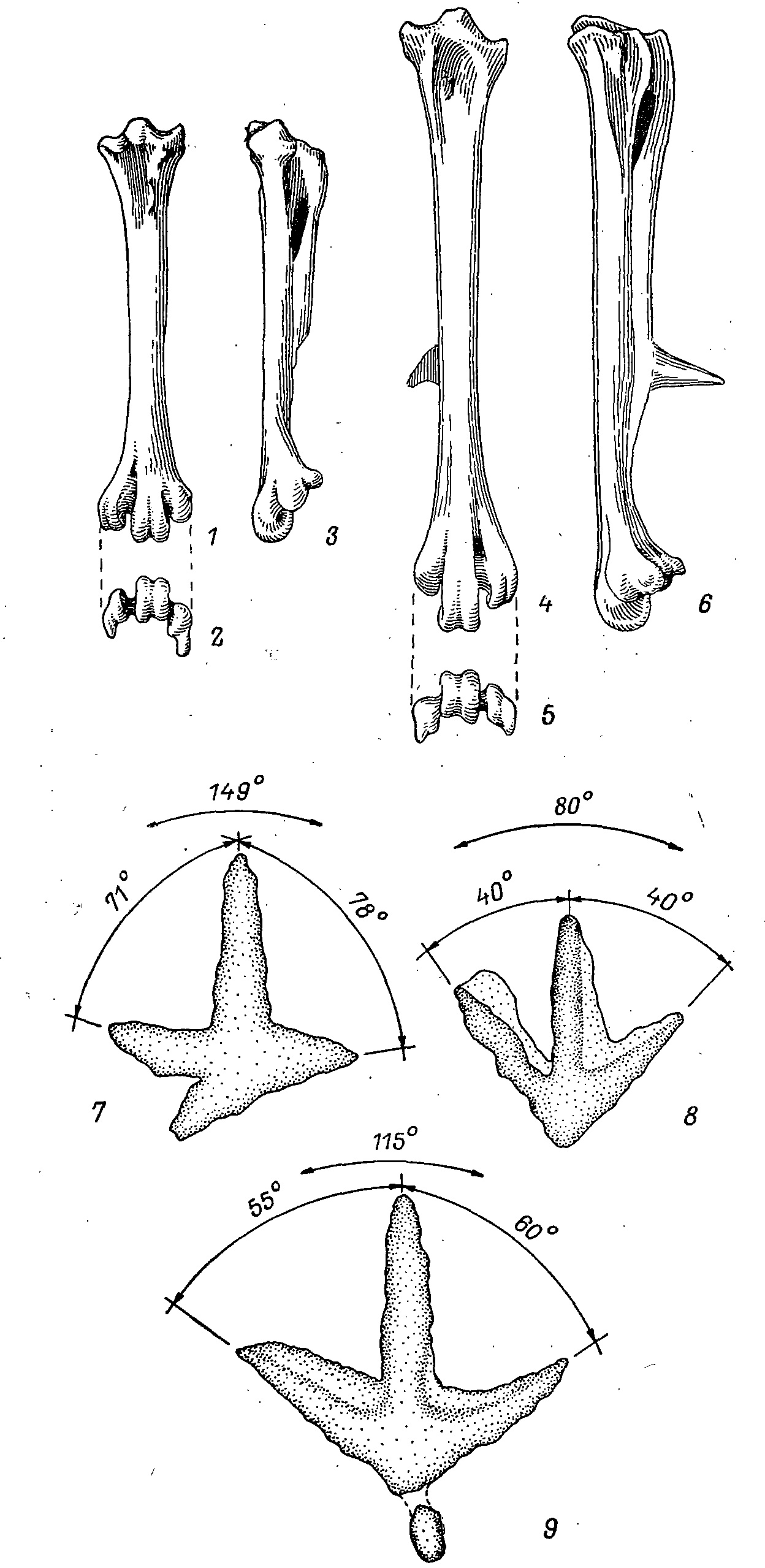
Рис. 5. Плюсневые кости (1—6) и расхождение пальцев (7—9) тетеревиных и фазановых птиц. 1—3 — Lyrurus tetrix, ♀; 4—6 — Phasianus colchicus, ♂ (1, 4 — вид спереди; 2, 5 — вид с дистального конца; 3, 6 — вид сбоку); 7 — след Lyrurus tetrix на снегу; 8 — след Tetraogallus himalayensis на снегу; 9 — след Phasianus colchicus на сыром грунте.
Основной особенностью скелета тетеревиных птиц является широкий и плоский таз. Он сильно уплощен в дорсовентральном направлении и сильно расширен в особенности в его каудальной части (табл. 1). Эта ширина обусловлена разрастанием боковых отростков позвонков в каудальной его части, а также резким расширением постацетабулярной части подвздошной кости (рис. 4).
Седалищные кости, наоборот, более узкие, чем у остальных куриных птиц, они ориентированы более наклонно по отношению к подвздошным костям в почти не вдаются под поверхность последних, так что сочленение этих костей по linea iliolateralis в отличие от остальных куриных имеет вид небольшого пологого гребня или едва намечается. Это и обусловливает небольшую высоту таза. Седалищные кости не выдаются далеко за задний край подвздошных костей, гребень crista transversus не столь высок и имеет более плоскую поверхность, a pr. pectinalis развит слабо. У большинства фазановых птиц последний отросток достигает большой величины и направлен рострально, хотя у отдельных видов (Ammoperdix griseogularis) он развит так же, как и у тетеревиных.
Большая ширина каудальной части подвздошной кости и пологая ориентация седалищной приводят к тому, что у большинства тетеревиных птиц (за исключением Lagopus lagopus и L. mutus) таз оказывается наиболее широким именно в этой области, тогда как у всех остальных куриных таз наиболее широк по линии выступов антитрохантеров (иногда ширина таза и здесь и там одинакова).
Изменения площади и конфигурации таза у тетеревиных птиц, казалось бы должны были повлечь за собой и изменения в мускулатуре задних конечностей, в особенности мышц, крепящихся к каудальной части седалищных костей, — m. ishio-femoralis и m. ileo-flexoreus. Однако их относительная величина осталась той же, хотя общая масса мускулатуры задней конечности тетеревиных по отношению к массе тела несколько ниже, чем у фазановых, — 7.7—10.5 % против 11.9—19 % (Кузьмина, 1977). Правда, данный автор использовал ограниченный набор видов, и можно ожидать некоторого перекрытия этих величин у двух семейств.
Особого внимания заслуживает заметно большее расхождение суставных бугорков пальцев на плюсне тетеревиных птиц, отмеченное Кузьминой (1964) и функционально объясненное мною (Потапов, 1974). Оно дает возможность широко расставлять пальцы, что особенно выгодно при ходьбе по снегу (140— 170° между 2-м и 4-м пальцами против 80—100° у фазановых птиц; рис. 5).
В мускулатуре наиболее примечательно отсутствие приводящей мышцы 2-го пальца, m. adductor digiti II, имеющейся у всех остальных куриных птиц (Hudson et al., 1959). Атрофия этого мускула могла произойти на стадии становления предковой формы семейства, и связано это было либо с древесным образом жизни, либо же с какими-то условиями функционирования лапы при снежном покрове.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
 Как кормить птенца и ухаживать за маленькой дикой птицей.
Как кормить птенца и ухаживать за маленькой дикой птицей. Пищеварительная система тетеревиных птиц.
Пищеварительная система тетеревиных птиц.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Подотека тетеревиных.
Подотека тетеревиных. Сезонные изменения окраски тетеревиных. Линька тетеревиных птиц.
Сезонные изменения окраски тетеревиных. Линька тетеревиных птиц. Фотоохота на кавказского тетерева в Тебердинском заповеднике.
Фотоохота на кавказского тетерева в Тебердинском заповеднике. Проблема смертности птиц от ЛЭП в Казахстане и способы защиты птиц.
Проблема смертности птиц от ЛЭП в Казахстане и способы защиты птиц. Окраска тетеревиных птиц.
Окраска тетеревиных птиц.
