Рамфотека тетеревиных птиц. Как птицы пережёвывают растения.
Для всех тетеревиных птиц характерна хорошо развитая рамфотека, образующая специфичный рельеф рогового нёба и формирующая режущие кромки челюстей. Предполагалось, что эта кромка позволяет работать челюстям по методу «ножниц» (Семенов-Тян-Шанский, 1959; Кузьмина, 1968; Дзержинский, 1977). Меня же прежде всего привлек рельеф рогового нёба. Было показано (Потапов, 1974), что нёбо у тетеревиных птиц ороговевает значительно сильнее, чем у большинства фазановых птиц, и имеет специфичный рельеф, состоящий из центрального и двух боковых валиков и желобка, идущего вдоль внутреннего края надклювья (рис. 1). Развитие этих валиков у разных видов варьирует. Они наименее выражены у Tetrao urogallus, имеющего наиболее плоское нёбо, и максимально развиты у Т. urogalloides, где получил исключительное развитие центральный валик, глубоко вдающийся в ротовую полость и предназначенный для дробления веток лиственницы — основного зимнего корма этого вида (Потапов, 1974). Все остальные виды тетеревиных птиц по степени выраженности валиков можно расположить в ряд между этими крайними вариантами, которые дают нам два представителя одного рода.

Falcipennis falcipennis - дикушка
В отношении самой механики отделения кормовых объектов (прочно связанных с растением концевых побегов, почек и хвои) существуют два представления. Согласно первому, этот корм состригается птицей по методу ножниц, где роль последних выполняют челюсти с режущей кромкой рамфотеки. По второму же, объекты отрываются от растений при помощи движений головы и шеи, а весь рельеф рамфотеки приспособлен только для жесткой фиксации объекта в клюве (Гамбарян, 1978). Я уже приводил ряд фактов (Потапов, 1974), показывающих невозможность рассматривать кормодобывающую деятельность всех тетеревиных птиц как «стрижку». Это особенно показывалось на примере Т. urogallus, который, по мнению ряда исследователей, обладает наиболее развитым механизмом типа ножниц (Дзержинский, 1974). Мною было показано, что главным у Т. urogallus при питании сосновой хвоей является подсекающее, движение головой, при котором захваченный клювом пучок хвои в основном отрывается. В таких случаях перерезались только концевые веточки, если они захватывались вместе с хвоей. В то же время я показал, что стрижка как таковая существует, в частности, у хвоеядной Falcipennis falcipennis, хотя кажется, что здесь имеет место не столько даже стрижка, сколько скусывание отдельных, хвоинок по методу «кусачек», описанного для вьюрковых птиц (Некрасов, 1964). В момент откусывания действует одна сторона клюва, правая, или левая, причем благодаря некоторому смещению нижней челюсти в соответствующую сторону режущие края надклювья и подклювья движутся навстречу друг другу в одной плоскости. Эта деталь не учитывалась авторами гипотезы ножниц, хотя любой вид тетеревиных, а, вероятно, и фазановых птиц при отрывании объекта никогда не режет его краями обеих челюстей сразу же с двух сторон. Объект всегда, срезается одной стороной клюва, и если, к примеру, птица поедает стебель травы или веточку дерева, то сначала скусывается верхушка, а потом птица продолжает откусывать кусочки с конца, никогда не стараясь выхватить кусок из середины (рис. 2). При этом как фазановая, так и тетеревиная птица старается ориентировать такую ветку или стебель не перпендикулярно к клюву, а под острым углом (Андреев, 1975б; Гамбарян, 1978; мои наблюдения). Важно учесть, что зимой при низких температурах ветки становятся хрупкими и при кормежке птица даже не стрижет и не кусает, а отламывает кусочки удобного размера. Хрупкость замерзших веток делает работу клюва исключительно эффективной, позволяя, например, каменному глухарю поедать до 360 кусочков в минуту без всяких дополнительных движений головы и шеи и тратить на эту деятельность не более 1.1 % суточного бюджета энергии (Андреев, 1975б). Однако при положительных температурах ветвь имеет высокую сопротивляемость на излом, и в таких случаях (так же, как и во всех случаях с хвоей) откусывание объекта усиливается подсеканием или подрезанием его кромками челюстей при помощи дополнительного рывка головы и шеи.
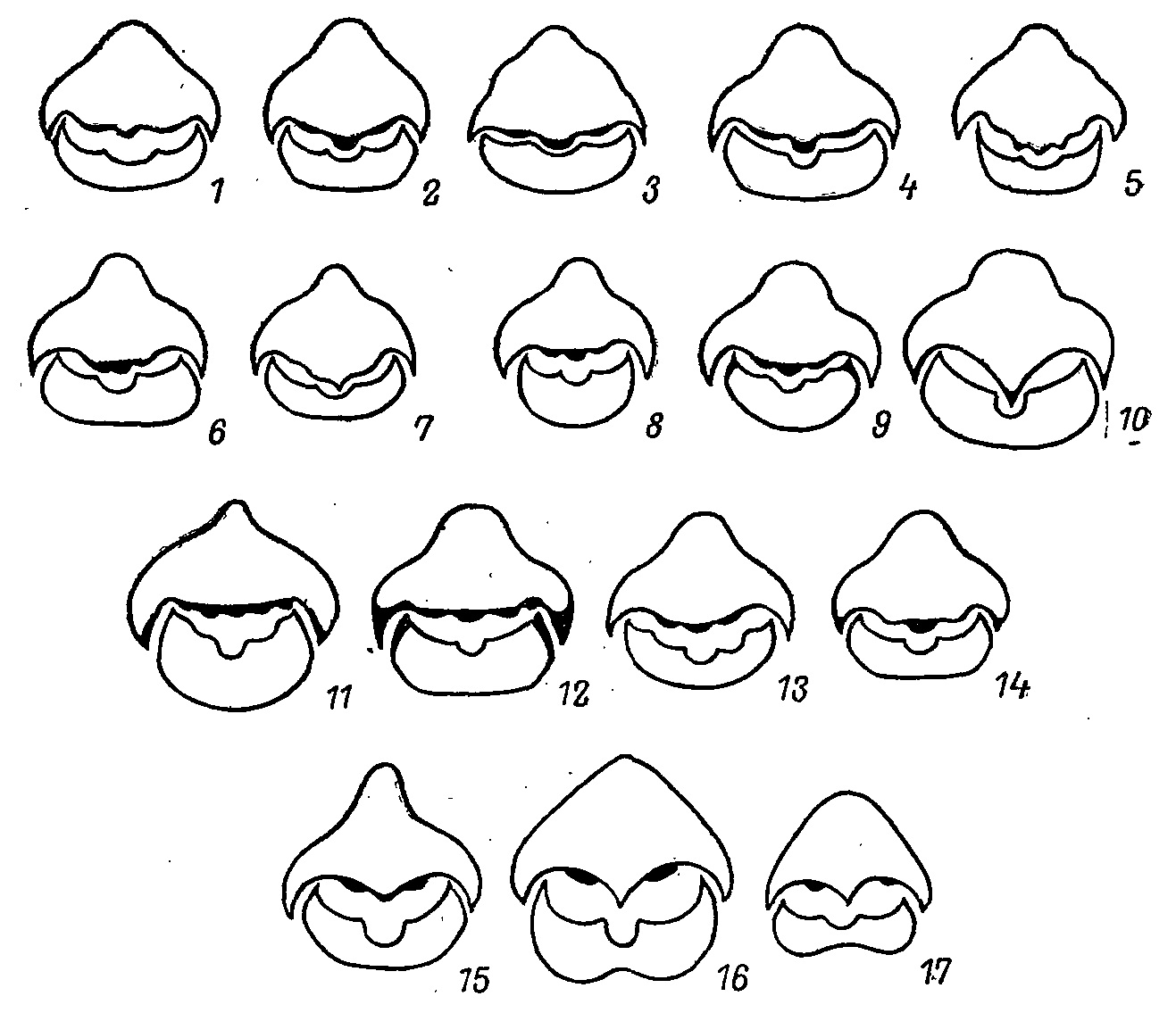
Рис. 1. Поперечные срезы клюва (на уровне заднего края ноздри). 1 — Bonasa bonasia; 2 — Falcipennis falcipennis; 3 — F. canadensis; 4 — Dendragapus obscurus; 5 — Lagopus mutus; 6 — L. lagopus; 7 — Lyrurus mlokosiewiczi; 8 — L. tetrix, ♂; 9 — L. tetrix, ♀; 10 — Tetrao urogalloides; 11 — T, urogallus, ♂; 12— T. urogallus, ♀; 13 — Bonasa umbellus; 14 — Tympanuchus phasianellus; 15 — Gallus gallus; 16 — Tetraogallus altaicus; 17 — Perdix perdix.
Необходимо отметить, что у фазановых птиц есть виды с очень сходным строением клюва вообще и рамфотеки в частности (Ithaginis). В зимнем питании видов рода Ithaginis важное место занимают концевые побеги пихт, сосен и можжевельника (Beebe, 1918; Ali, Ripley, 1969). Поэтому, оценивая различия фазановых и тетеревиных птиц в строении рамфотеки и способах поедания корма, приходится признать, что различия эти скорее количественного характера. Как и в ряде других случаев, сем. Phasianidae показывает в этом отношении значительно более широкий спектр вариаций, чем более узко специализированные тетеревиные птицы.
К сожалению, неизвестно, как протекает линька рамфотеки у тех видов фазановых, у которых строение клюва сходно с таковым у тетеревиных. У фазановых же таких родов, как Phasianus, Tetraogallus, Alectoris, рамфотека растет постоянно по мере снашивания (Кузьмина, 1968). У тетеревиных птиц линька рамфотеки носит четко выраженный сезонный характер (Снигиревский, 1950) и наиболее интенсивно, протекает в июле, когда потребление древесных кормов почти полностью прекращается. Заканчивается линька в сентябре, и рамфотека полностью обновляется к началу зимнего сезона, в котором она несет максимальную нагрузку.
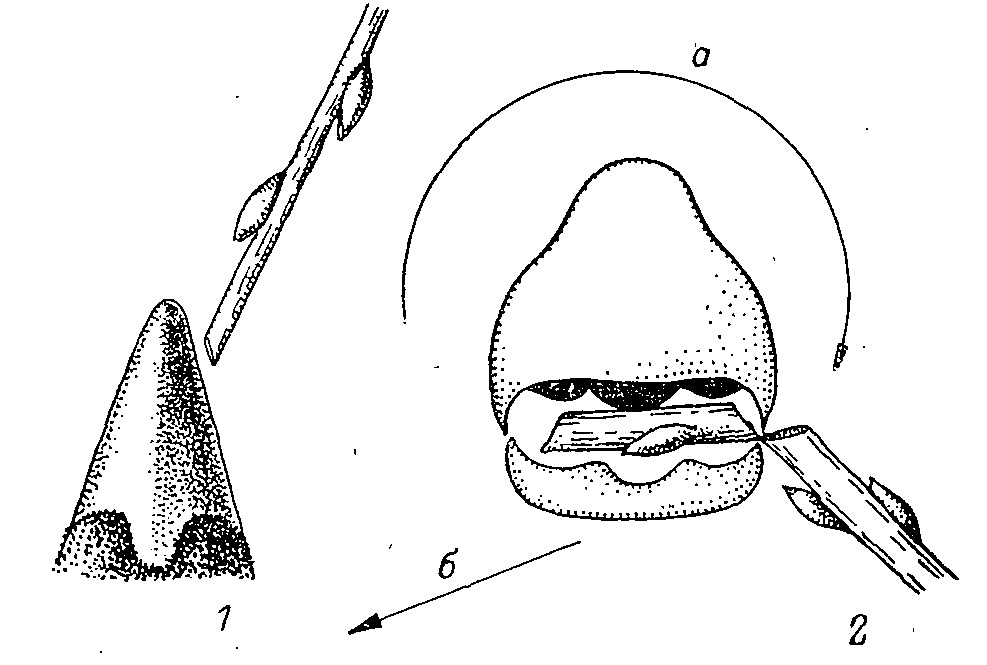
Рис. 2. Схема скусывания побега тетеревиной птицей. 1 — ориентация клюва по отношению к поедаемому побегу; 2 — положение побега в клюве при скусывании. а — направление вращательного (подсекающего) движения; б — одновременного рывка головы в сторону.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные."
 Пищеварительная система тетеревиных птиц.
Пищеварительная система тетеревиных птиц. Зоб тетеревиных птиц. Сколько еды может вместить зоб?
Зоб тетеревиных птиц. Сколько еды может вместить зоб?Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Скелет и мускулатура тетеревиных птиц.
Скелет и мускулатура тетеревиных птиц. Как кормить птенца и ухаживать за маленькой дикой птицей.
Как кормить птенца и ухаживать за маленькой дикой птицей. Подотека тетеревиных.
Подотека тетеревиных. Сезонные изменения окраски тетеревиных. Линька тетеревиных птиц.
Сезонные изменения окраски тетеревиных. Линька тетеревиных птиц. Фотоохота на кавказского тетерева в Тебердинском заповеднике.
Фотоохота на кавказского тетерева в Тебердинском заповеднике.
