Азиатская дикуша. Falcipennis falcipennis.
Лесная птица средних размеров, несколько крупнее обыкновенного рябчика. Крыло сильно закруглено, вершину его у самцов образуют 7-е, реже 8-е или 6-е и 7-е, или даже 6-е, 7-е и 8-е ПМП (первостепенные маховые перья) вместе, а у самок 7-е, 7-е и 8-е или реже 6-е и 7-е ПМП. Такие вариации в форме верхушки крыла объясняются сильной заостренностью дистальных ПМП, которые в различной степени подвержены износу. Резкая заостренность дистальных маховых перьев, имеющих к тому же серповидную форму, отличает азиатскую дикушу от всех тетеревиных птиц и в свое время послужила поводом для выделения вида в монотипичный род. И у самцов, и у самок наиболее заострены три самых дистальных маховых пера, затем заостренность быстро уменьшается, сходя на нет уже на ПМП-5, а четыре первых ПМП имеют нормальную форму. Такое резкое заострение ПМП и их изогнутая форма привели к заметному укорочению длины крыла, и азиатская дикуша — наиболее короткокрылый вид в семействе (отношение длины крыла к основному размеру — 175.2 %). Наиболее сильно перья заострены у старых самцов. Плюсна оперена до основания, основная фаланга 3-го пальца оперена на 2/3 длины, у 2-го и 4-го пальцев оперено самое основание первой фаланги со стороны перепонки. Когти довольно длинные, заметно изогнуты. Оперение плюсны, отрастающее к началу зимы, очень густое и скрывает пальцы до самых когтей. Половой диморфизм хорошо выражен в окраске оперения и почти незаметен в размерах.

Самец азиатской дикуши.
Образ жизни.
Питание.
Оперение и окраска.
Размеры и масса.
Линька.
Ареал и распространение.
Места обитания.
Численность.
Территориальность.
Размножение и токование.
Гнездование.
Хозяйственное значение и охрана.
Образ жизни.

Самка дикуши азиатской.
Наиболее специфичная черта в поведении как азиатской, так и американских дикуш — их чрезвычайная «доверчивость», отсутствие боязни человека и даже собак. При появлении человека птицы не улетают, а либо не спеша удаляются пешком, либо взлетают на ближайшее дерево, где спокойно сидят и не улетают ни при выстрелах, ни при попытках, нередко успешных, поймать их петлей, надетой на палку. Птицы, расхаживающие по земле, подпускают наблюдателя почти вплотную и позволяют длительное время следить за собой, нередко не обращая на него никакого внимания. Причину такого поведения никак нельзя объяснить «тупостью», неспособностью реагировать на опасность, как то делал Л. М. Шульпин, — ведь это свойство всех видов рода, живущих на различных материках. Правильную, на наш взгляд, точку зрения высказал в свое время Афанасьев (1934), который считал «бесстрашие» дикуш защитным приспособлением. Действительно, подробные наблюдения как наших, так и американских исследователей показали, что видимое отсутствие боязни человека сочетается с еще одной очень характерной чертой — малой подвижностью. Кормящаяся на дереве или на земле птица то и дело надолго замирает, как бы дремлет. Движения их медленные, плавные, они редко пользуются полетом. И, тем не менее, такая «заторможенность» птиц — кажущаяся. Дикуши все время, даже при дремоте, пребывают настороже, постоянно прислушиваются, причем слух у них судя по всему очень хороший. Особенно красноречивы факты, показывающие результаты такого поведения. Так, в одном случае самка дикуши, обитавшая с тремя подросшими птенцами в непосредственной близости от логова соболя, ни разу от него не пострадала. В этой же ситуации собака-лайка, бывшая у исследователей, неоднократно ловила молодых тетеревов и глухарей, но ей ни разу не удалось поймать ни взрослых, ни птенцов дикуши — во всех случаях птицы успевали взлетать на деревья. Незаметности птиц способствует также покровительственная окраска, особенно эффективная зимой, а также крайняя молчаливость. Даже токовое поведение у них очень тихое, наименее заметное среди всех лесных тетеревиных птиц. О звуках, издаваемых самцом вне токования, вообще ничего неизвестно — указаны только тихие скрежещущие звуки, издаваемые самкой, вспугнутой собакой, вперемежку с тихим клохтанием, и тихие позывки типа «куть-куть-куть», также издаваемые самкой, вспугнутой в паре с самцом (Абрамов, 1962). Есть указания на то, что самка при тревоге издает грубое квохтание, как тетерка, но менее громкое (Капланов, 1938).

Рис. 1. Азиатская дикуша Falcipennis falcipennis. 1 — самец; 2 — самка.
Все виды дикуш отдают явное предпочтение передвижению пешком, а не на крыльях. Л. М. Шульпиным описан случай, когда дикуша пыталась уходить пешком даже от собаки.
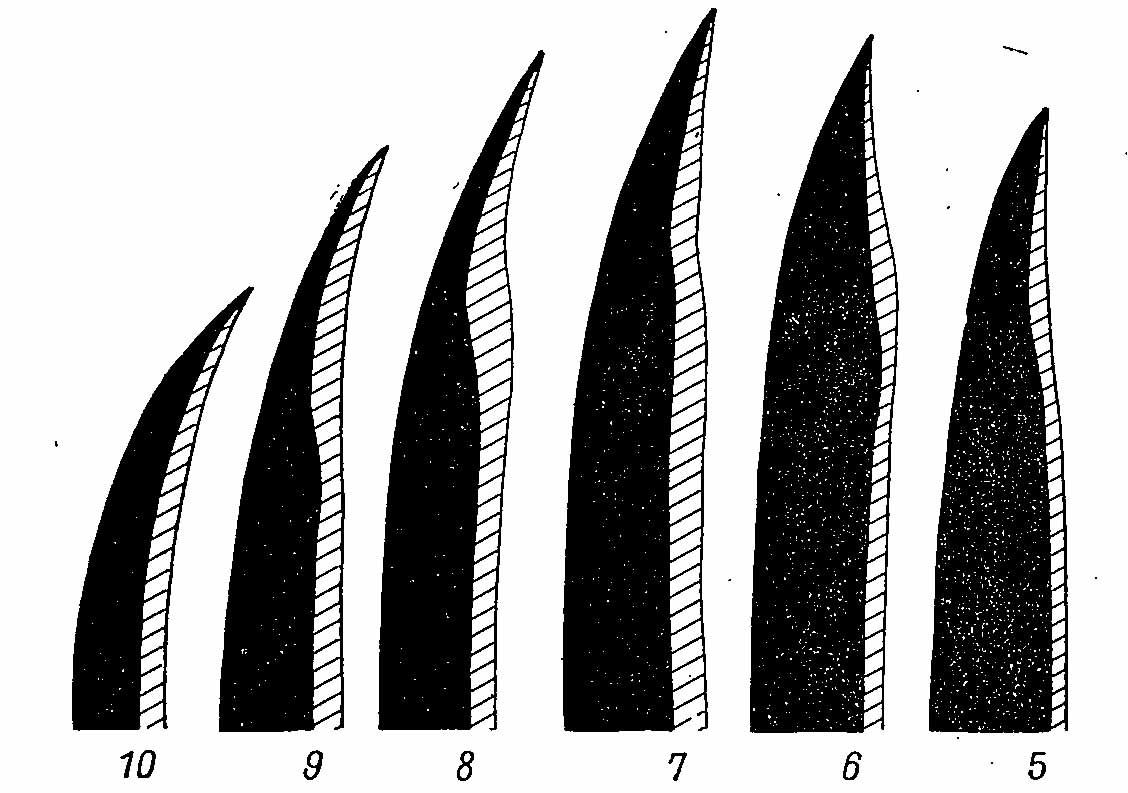
Рис. 2. Форма дистальных первостепенных маховых перьев самца Falcipennis falcipennis. В контур перьев годовалой птицы (заштриховано) вписан контур перьев взрослой птицы (сплошная заливка). Цифры указывают номера первостепенных маховых перьев.
Полет дикуш — обычного для тетеревиных птиц типа. Самцы и самки, будучи внезапно вспугнутыми, взлетают вверх с большим шумом и видимыми усилиями, громко хлопая крыльями и задевая ими за ветки. Возможно, что при других ситуациях взлет птиц может быть легким и бесшумным. Мне приходилось при ловле токовавшего самца наблюдать, как он, избегая моего преследования, сравнительно легко взлетал вверх на быстро вибрирующих крыльях, издававших при этом легкий жужжащий звук, и перемещался на несколько метров в сторону в довольно густых зарослях, умудряясь не задевать при этом ни одной ветки. По данным А. С. Никанорова (устное сообщ.), полет дикуши сопровождается специфичным свистом крыльев, а дальность его обычно не превышает 20-30 м. Максимальная дальность перемещения — 100 м — была отмечена при перелете на ночевку, причем перед таким перелетом птицы, перескакивая с ветки на ветку, поднимались для старта на самую вершину дерева.
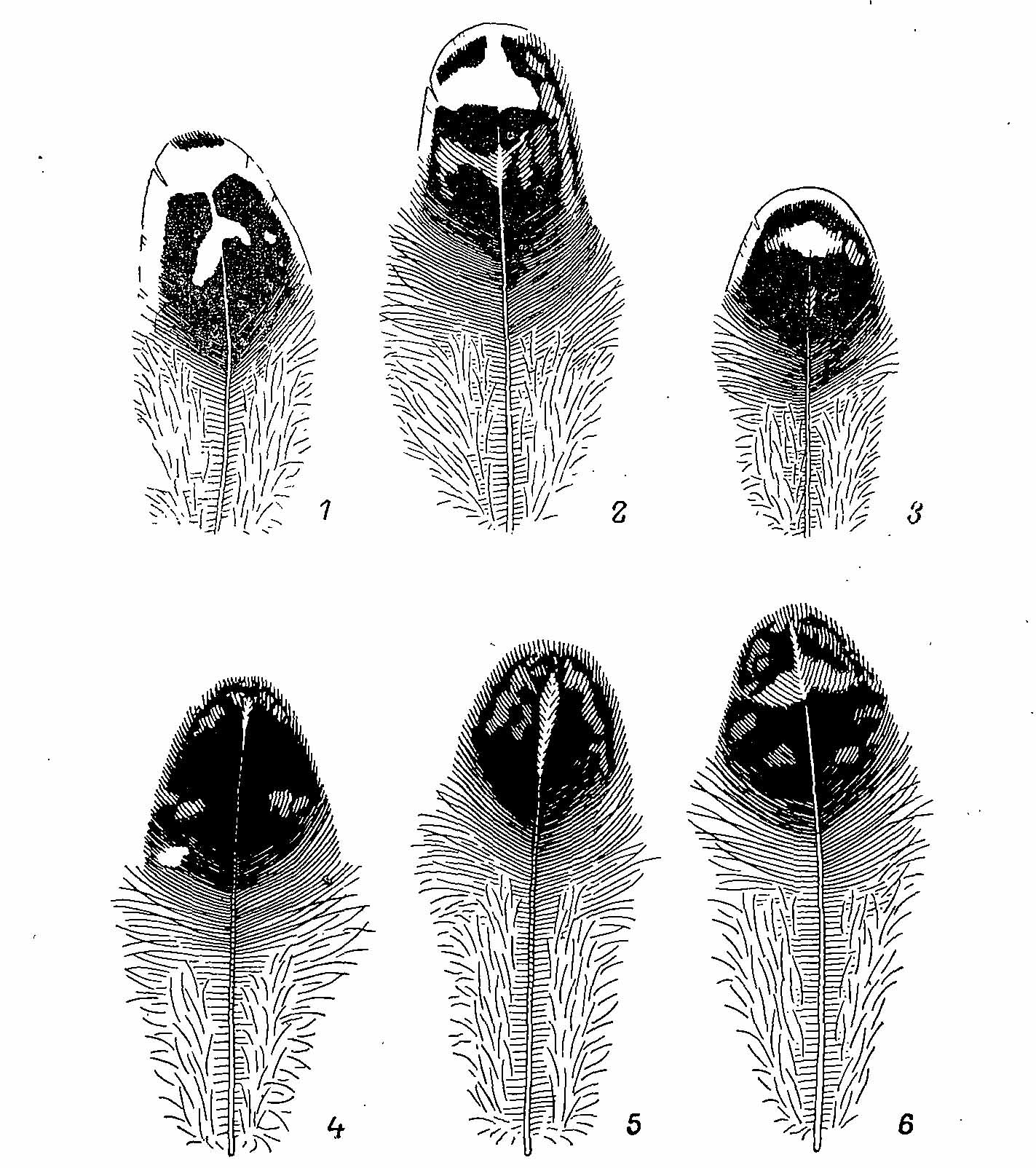
Рис. 3. Перья самки Falcipennis falcipennis. 1 — с груди; 2 — с боков тела; 3 — с передней части брюха; 4 — со спины; 5,6 — с поясницы (два типа окраски).
В заключение следует отметить, что эта своеобразнейшая черта в поведении дикуш делает их исключительно перспективным объектом для исследования самых тонких черт биологии — территориальности, динамики популяционного состава и численности, питания, размножения и т. п. Американские исследователи уже используют эти возможности в полной мере.
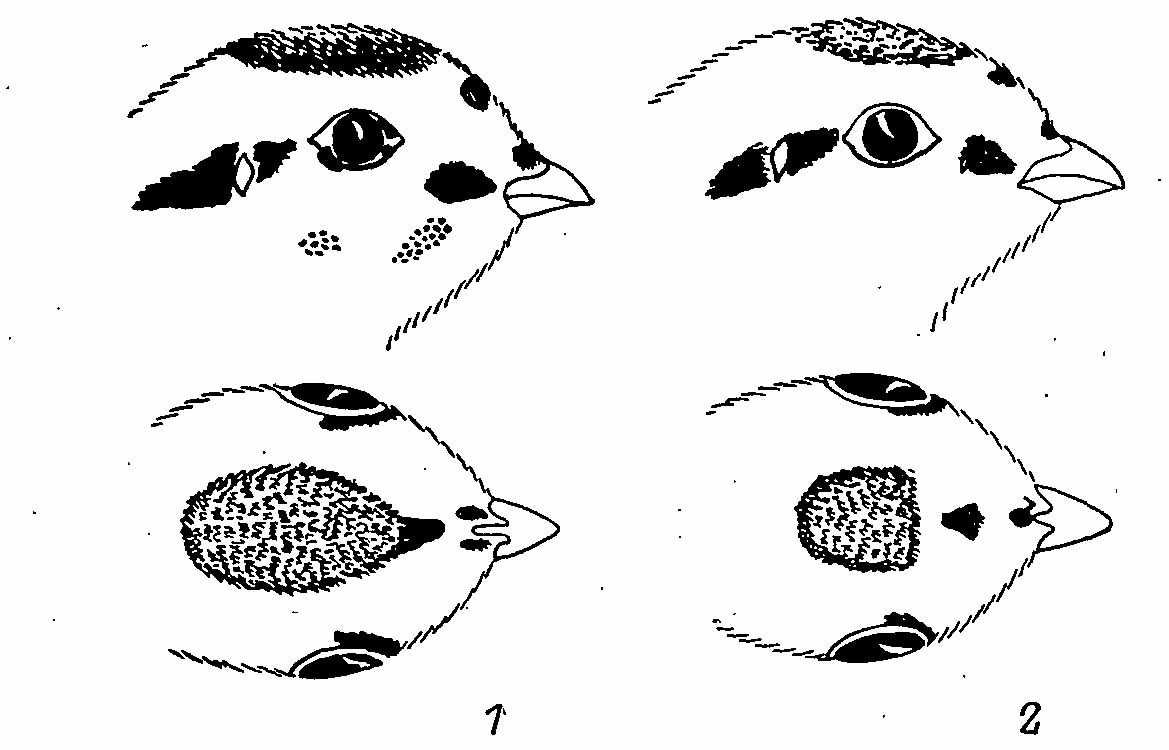
Рис. 4. Рисунок на голове пухового птенца Falcipennis falcipennis (1) и F. canadensis (2).
Питание.

Таблица 1. Размеры Falcipennis falcipennis
Зимнее питание состоит почти исключительно из хвои пихты и елей, но, видимо, из-за трудности определения видовой принадлежности хвои данные на этот счет весьма противоречивы. Так, для Сахалина основным зимним кормом указана хвоя пихты (Мишин, 1959). В бассейне р. Селемджи хвоя белокорой пихты является основным кормом с августа по май; в меньшей степени поедается хвоя аянской ели (Юдаков, 1972). В питании молодых птиц этот корм был отмечен с 18 августа. В районе оз. Кизи в качестве основного зимнего корма также указана хвоя пихты (Воробьев, 1954). В то же время А. С. Никаноров (устное сообщ.) до 10 мая в верховьях р. Бикин находил в зобах почти исключительно хвою аянской ели. В Якутии дикуши питаются зимой хвоей аянской и обыкновенной елей, используя в качестве добавки верхушки хвоща, торчащего из-под снега (Воробьев, 1963). Наблюдения А. Г. Юдакова и отснятые им фотографии показывают, что птицы не отрывают хвоинки, а отстригают их, сопровождая эту операцию характерным вращательным движением головы (согласно А. С. Никанорову — с резким поворотом головы). Основание хвоинки остается на ветке и имеет довольно ровный край среза. По-видимому, основания хвоинок оставляются на ветвях из-за их большой смолистости.
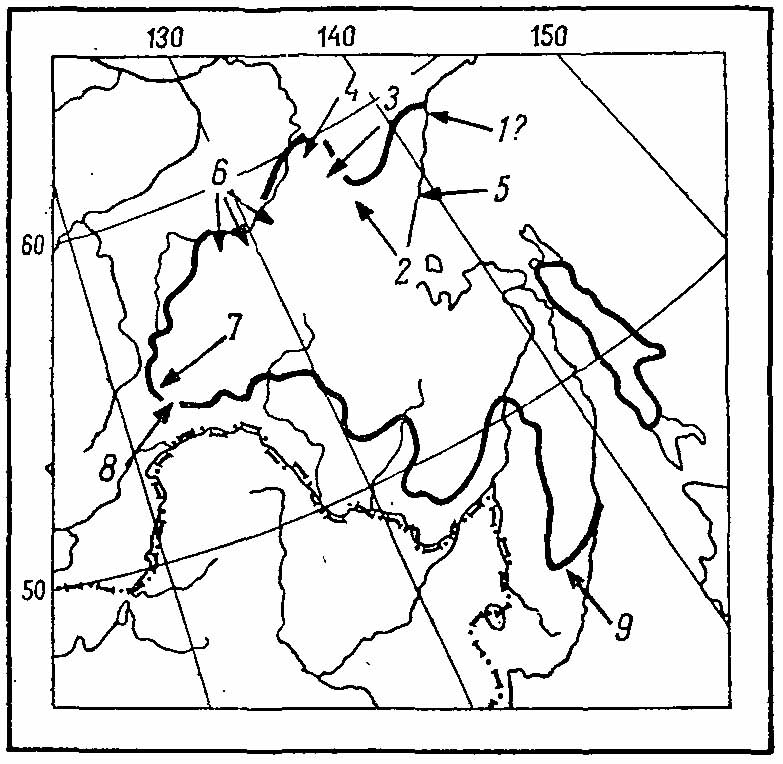
Рис. 5. Ареал Falcipennis falcipennis. 1 — р. Кухтуй у Охотска (Р. Л. Потапов, расспросные данные), Охотск (Воробьев, 1954); 2 — Нелькан (кол. ЗИН АН СССР); 3 — р. Ивгили (Перфильев, 1975); 4 — низовья р. Маи (Перфильев, 1975); 5 — Аян (кол. ЗИН АН СССР); 6 — Алдан (правые притоки, О. В. Егоров, in litt.); 7 — верховья р. Нюкши (А. Я. Тугаринов, in litt.); 8 — верховья р. Ольдой (Гассовский, 1927); 9 — верховья р. Рудная (Шульпин, 1936).
Весной в корме продолжает преобладать хвоя аянской ели, к которой прибавляются распускающиеся почки хвои лиственницы и реже корейского кедра, а также начинают поедаться верхушки мха (Pleurosium schreberi) (неопубликованные данные Р. Л. Потапова и А. С. Никанорова). По данным последнего автора, весь июнь и начало июля дикуши питаются преимущественно верхушками мха и листьями брусники, в меньшей степени — хвоей аянской ели и лиственницы. Ни разу не было замечено питание хвоей белокорой пихты или сибирской ели. Все авторы отмечают в составе летнего корма хвою лиственницы и пихты, ягоды морошки, клюквы, вороники, листья брусники (Гизенко, 1955), а у молодых птиц — листья и ягоды брусники, голубики, муравьев, кузнечиков и жуков (Абрамов, 1962), семена осок и вороники (Афанасьев, 1934). Осенью наряду с хвоей лиственницы и пихты в большом количестве поедаются различные ягоды (черника, голубика, брусника, морошка, клюква, вороника, малина), семена осок, костяники и в очень небольшом количестве — насекомые. Таким образом, хвоя, составляя основной корм в зимнее время, продолжает оставаться важным кормовым объектом в течение всего теплого времени года.

Рис. 6. Песенный ритуал Falcipennis falcipennis. 1 — расхаживание; 2 — стартовая поза; 3 — движение хвоста в момент остановки; 4 — «урчание»; 5 — двойной прыжок со щелчками и разворотом.
По данным А. С. Никанорова, в питании птенцов очень мала доля насекомых. В вольерах они уже с 2-недельного возраста обрывают хвоинки ели и лиственницы, которые к месячному возрасту уже занимают видное место в их рационе.
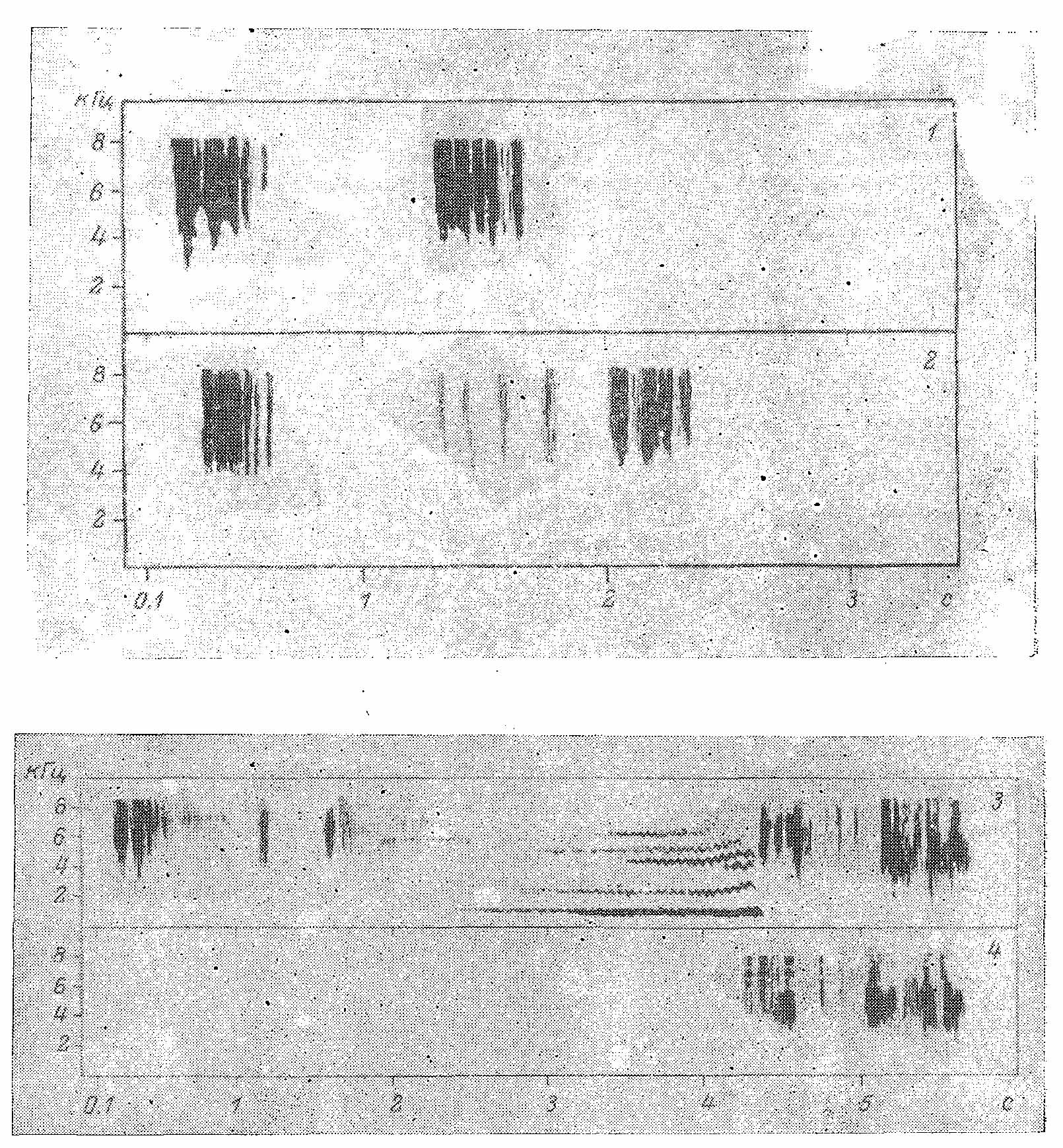
Рис. 7. Сонограмма звуков при раскрывании и складывании хвоста (1, 2) и полного песенного ритуала (3,4) (по: H. Bergmann, in litt.). На сонограмме отчетливо видно последнее перед пением движение хвоста, само вибрирующее пение (волнообразные линии) и сгущения шумов, соответствующие первому одиночному и двум двойным щелчкам. По оси ординат — уровень звука, кГц; по оси абсцисс — время, в с.
Зимний сезон. Сведений о зимней жизни очень мало. Всю зиму птицы держатся небольшими стаями (иногда до 30 особей; Юдаков, 1972), в это время они максимально оседлы и проводят весь зимний сезон на крайне ограниченной площади. Максимальное количество корма в зобу (хвоя белокорой пихты) — 39 г (сухая масса). В данном случае птица была добыта 25 ноября после захода солнца.
Оперение и окраска.
Взрослый самец.
Голова черновато-серая, перья здесь черноватые с серым окаймлением и с буроватыми поперечными полосками, обычно прикрытыми налегающими перьями. На перьях зашейка каймы и полосы шире и зашеек кажется более светлым, чем голова. Иногда широкие серые каймы почти совсем скрывают бурый цвет перьев и зашеек выглядит сплошь серым. Межлопаточная область имеет наиболее темную окраску — перья здесь черные с узкими светло-серыми каймами, создающими чешуйчатый рисунок. Задняя часть спины несет струйчатый рисунок, образуемый неровными поперечными серовато-охристыми полосами. На пояснице эта струйчатость становится более интенсивной, появляются охристые наствольные полоски, которые ближе к хвосту становятся очень четкими и приобретают белый цвет. ВКХ (верхние кроющие хвоста) окрашены так же, но белых наствольных полос не имеют. На боках головы перья уздечки своими белыми вершинами формируют белую полосу, идущую к глазу и продолжающуюся за ним. Ушные перья черно-бурые, с неясным поперечным рисунком из темно-серых полос. Горло черное, с небольшим количеством белых пятнышек и окаймлено неширокой белой полосой, которую формируют широкие предвершинные полосы перьев. Вслед за белым окаймлением горла следует пестро окрашенный участок шеи, где перья несут поперечный рисунок из охристых полос по черновато-бурому фону. Грудь окрашена в черно-бурый цвет (у живых птиц, особенно у старых самцов она выглядит совершенно черной, но быстро буреет в процессе хранения). В нижней части груди окраска становится несколько светлее, на перьях появляется поперечный рисунок из волнистых белых предвершинных полос и узких беловатых каемок. На перьях боков груди эти полосы принимают форму наствольных пятен, а на перьях брюха они становятся более широкими, принимают полукруглую, а ближе к хвосту и на боках — треугольную «сердцевидную» форму. На многих перьях брюха и боков тела эти пятна прорывают черный бордюр и выходят к вершине пера. Особенно правильный сердцевидный рисунок несут перья на боках тела. На перьях, расположенных ближе к подхвостью, ниже упомянутого белого пятна есть еще одна белая полоса. Почти на всех перьях нижней части тела, несущих характерные белые пятна, есть еще неясные струйчатые полосы охристого или серого цветов — они обычно скрыты налегающими вершинами соседних перьев. Перья подхвостья с широкими белыми вершинами и неясным поперечным рисунком серовато-желтого цвета по черному фону. ПМП черно-бурые, самые внутренние — с беловатыми кончиками. ВМП несколько светлее, с тонкой беловатой каймой и легким струйчатым рисунком по наружному краю опахала (рисунок этот может полностью отсутствовать). На самых внутренних ВМП серый струйчатый рисунок хорошо развит, особенно на внешнем опахале. На этих же перьях есть наствольные белые полосы. Большие ВКК (верхние кроющие крыла) однообразной черно-бурой окраски, средние и малые ВКК такого же цвета, но с широкими серовато-бурыми каймами и серым струйчатым рисунком. На проксимальных партиях этих перьев есть белые наствольные полоски и пятна, расположенные у вершин перьев; число и величина их варьируют индивидуально. На черно-шоколадном фоне плечевых перьев, особенно на наружном опахале, хорошо развит серый или серовато-желтый струйчатый рисунок, с крупными предвершинными наствольными пятнами белого цвета. Они наиболее развиты на самых длинных плечевых перьях и, расширяясь, выходят к вершине пера. Подмышечные перья — с белыми вершинными каймами и поперечными полосами по шоколадному фону. РП (рулевые перья) черные с широкими белыми вершинными пятнами, за исключением центральной пары, где пятно это очень маленькое. На этой же паре перьев есть легкий струйчатый рисунок в верхней части пера. Птица в свежем пере выглядит более желтовато-охристой.
Самец в первом взрослом наряде отличается от старого менее заостренными ПМП (рис. 2) и незначительными деталями в окраске. Наствольные полоски на перьях поясницы не такие заметные и почти всегда не белого, а желтоватого цвета. Темный пластрон на груди светлее и значительно уже, чем у старых птиц. Центральные РП с большим развитием струйчатого рисунка и либо без вершинного белого пятна, либо оно едва выражено. 2-3 пары боковых РП несут на внешних опахалах остатки поперечно-полосатого рисунка. НКХ (нижние кроющие хвоста) более светлые, с хорошо развитыми поперечными полосами.
Взрослая самка.
Общая окраска верха пестрая, буровато-рыжая. Перья головы и шеи с охристыми и серыми поперечными полосами. На перьях спины помимо этих полос есть еще наствольные продольные пятна, особенно четко выраженные на пояснице и надхвостье. В окраске самок, как и у многих других тетеревиных птиц, наблюдается дихроматизм в виде двух вариантов окраски — серой и охристой. Это зависит от преобладания того или иного цвета в окраске полос и пятен. Однако помимо цветовых вариантов у самок азиатской дикуши имеются два типа рисунка на перьях поясницы (рис. 3) причем эти типы рисунка никак не связаны с возрастными изменениями и встречаются примерно с равной частотой. Горло пестрое, беловато-охристое. На шее и зобу преобладает охристая окраска, создаваемая поперечными полосами на перьях, общий тон которых бурый. На фоне общей беловатой окраски низа область зоба выделяется поэтому широкой охристой полосой. Нижняя часть груди, брюхо и бока тела окрашены, как у самцов, но белые пятна и полосы развиты значительно сильнее и, налегая друг на друга, часто образуют сплошные белые поля. Окраска крыла и хвоста такая же, как у самцов, на них в верхней части перьев развиты поперечные охристые полосы, хотя белые вершины остаются такой же величины, как и у самцов. Летом светлые каймы перьев обнашиваются и окраска становится более темной, в особенности на голове и в межлопаточной области.
Ювенальный наряд (самец и самка).
Верхняя часть тела пестрая, из охристых поперечных полос и бурых поперечных полос и пятен. Кроющие и плечевые перья крыла с охристыми стержневыми полосами и белыми вершинными пятнами. ПМП бурые с беловатыми каймами по наружному опахалу и с рыжеватыми крапинками на внешних опахалах самых внутренних маховых. На ВМП — поперечные желтоватые и бурые полосы, на самых внутренних перьях эти полосы черного и бурого цвета, резко выражены и пронизаны наствольной охристой полосой. РП бурые, с желтоватыми и белесыми поперечными полосками. Горло белое, окраска груди охристого оттенка, брюхо беловатое. На боках тела перья с рисунком из белых и охристых полос.
Пуховой наряд.
Нижняя часть тела бледного палево-желтого цвета, резко контрастирующего с яркой желтовато-коричневой окраской верха. На темени — коричневая шапочка с едва намеченным черным окаймлением. Это окаймление меньше выражено, чем у птенцов американских дикуш, и иногда может совсем отсутствовать (из 16 пуховых птенцов с о-ва Сахалин в Орнитологическом институте Ямасины в Токио это окаймление отсутствовало у 1 птенца). Окраска пуховых птенцов с верховьев р. Большая Уссурка (сборы Ю. Б. Пукинского) не отличается от таковой сахалинских. На боках головы имеется узор из черных пятен (рис. 4), причем лобовое пятно обычно соединяется с черным окаймлением шапочки, но у некоторых экземпляров этого соединения нет. Две черные отметинки на щеках имеются примерно у 50% особей.
Размеры и масса.
Помимо коллекции ЗИН АН СССР использовано 32 экз. с южного Сахалина из коллекции Орнитологического института Ямасимы (Токио).
Размеры самцов (34 штуки): крыло 182.7 мм (174-195 мм),. хвост 107 (98-121), клюв 10.7 (8.3-12), плюсна 36.8 (33-37.5), средний палец 36 (34-39.2).
Самки (25): крыло 184.1 мм (175-196 мм), хвост 102 (95-106), клюв 10.7 (9-12.4), плюсна 35 (32-38), средний палец 35.3 (32-38).
Птицы с Сахалина, вроде бы, отличаются несколько большей длиной крыла и среднего пальца (табл. 1), но материала для выделения этих птиц в отдельный подвид только на этом основании явно недостаточно. Необходимо иметь в виду, что по крайней мере у самцов с возрастом обычно (но не всегда) увеличивается длина хвоста и крыла. Если годовалые самцы имеют в большинстве длину хвоста 101-107 мм, то у взрослых она 107-115 мм.
Данных по массе очень мало. Взрослый самец, добытый мной в низовьях Амура 11 мая, имел массу 736 г, добытая рядом с ним самка с развитым яичником — 730 г. Молодой самец, добытый здесь же двумя днями раньше, имел массу 662 г. Масса двух самцов, добытых в июле и августе, была соответственно 600 и 640 г (Абрамов, 1962). Для молодого самца с Алдана (октябрь) на этикетке указана масса в 635 г (кол. ЗИН АН СССР). Средняя масса самцов и самок с Селемджи, без разделения по сезонам и по возрасту птиц, соответственно 708 и 636 г (Юдаков, 1972). Эти данные говорят о существенном различии в массе годовалых и взрослых птиц (по, крайней мере самцов) — разница достигает 100 г.
Линька.
Линька протекает по схеме, общей для тетеревиных птиц. Из особенностей можно отметить только почти полное отсутствие перьев летнего наряда. Сроки же таковы. Молодые птицы заканчивают линьку в первый взрослый наряд к середине сентября. Когда начинают линять взрослые самцы — неизвестно, майские экземпляры (10 мая, р. Учур, 10 и 11 мая, Богородское, р. Амур, кол. ЗИН АН СССР) линьку еще не начали. Самец от 29 июня (верховья р. Большая Уссурка, кол. ЗИН АН СССР) — в обношенном оперении, сменились только 5 дистальных ПМП, летних перьев нет. Самец от 2 августа (бассейн р. Сутана, кол. МГУ) почти весь перелинял, дорастают рулевые, плечевые и контурные перья головы. Взрослый самец с верховьев р. Горин от 31 июля сменил все маховые перья, рулевые — в пеньках, долинивает контурное оперение на шее и спине (Абрамов, 1962) — данные эти вызывают сомнение. Самец от 3 августа с зал. Де-Кастри (ЗИН АН СССР) меняет ПМП-8, из второстепенных сменились 2-5-е и 13-14-е, дорастают РП, старое оперение сохранилось главным образом на голове, шее, груди и плечах. Наряду с этим самец от 4 августа с Сахалина сменил только 5 ПМП, РП еще старые, интенсивная линька контурного оперения еще продолжается. Самец от 3 сентября с мыса Лантарь (Охотское побережье) заканчивает линьку — дорастают ПМП-8, ПМП-9 и ПМП-10, целый ряд контурных, плечевые же перья только вышли из кисточек. Экземпляр от 7 сентября из-под Николаевска практически перелинял — дорастают отдельные контурные перья на голове, груди и шее. У самцов от 14 сентября (Немуй, Охотское побережье) и от 26 сентября (Аян) линька также заканчивается, дорастают отдельные перья на брюхе. Самец от 14 октября из хр. Джагды тоже заканчивает линьку — есть чехлики на основаниях брюшных перьев, дорастают плечевые. Экземпляр от 6 октября с Учура полностью закончил линьку. В целом можно считать, что наиболее интенсивно линька у большинства особей протекает в июле и к середине августа основная масса перьев бывает уже сменена. Однако же долинивание может тянуться еще долго, до середины октября.
Самки начинают линять гораздо позднее. Летних перьев у них тоже пока не обнаружено, хотя некоторое разреживание оперения явно происходит. Сама линька начинается, видимо, сразу же после окончания насиживания. Самка от 11 июля со Станового хребта с начавшим зарастать наседным пятном сбросила только ПМП-1. Самки от 23 июля (мыс Лантарь, Охотское побережье) и от 26 июля (Туксаин, Становой хребет) меняют ПМП-3, наседное пятно зарастает, но контурное оперение почти сплошь старое. У самок от 23 и 26 июля с верховьев Амгуни сменяется ПМП-4, наседное пятно зарастает, но контурное оперение старое. У самки с верховьев Амгуни от 2 августа дорастает ПМП-4, зарастает наседное пятно, преобладает старое контурное оперение, но всюду под ним, кроме головы, растут новые перья; у самки с тех же мест от 7 августа сменилось ПМП-5, но среди контурных перьев преобладают старые. У самки от 13 августа со Станового хребта сменилось 6 ПМП, началась смена РП, новые контурные перья всюду еще скрыты под старыми, наседное пятно только начало зарастать. Самка от 28 августа с Бурей сменила ПМП-6, старые РП выпали, идет бурная линька контурного оперения, но старые перья еще преобладают. У самки от 24 августа с оз. Кизи (Зоомузей МГУ) РП отрасли на 1/3 длины, дорастает ПМП-6, новые контурные перья преобладают только по бокам, на спине же они еще скрыты старыми. У самки с Сахалина от 15 сентября (Зоомузей МГУ) долинивает контурное оперение, дорастают ПМП-9 и ПМП-10. Видимо, в этом месяце линька у самок заканчивается. Судя по состоянию оперения у самки от 13 августа, явно имевшей очень поздний выводок, линька началась задолго до вылупления птенцов. Видимо, насиживание сдерживает начало линьки только до определенного момента.
Ареал. Распространение.
Область обитания азиатской дикуши в целом невелика и охватывает дальневосточную тайгу охотского типа к югу от 60° с. ш. (рис. 5). Распространение дикуши, как и ее основного зимнего корма, аянской ели (Picea jezoensis), крайне мозаично: достоверных сведений о границах ареала очень мало. В целом к настоящему времени они определяются следующим образом. На север вид идет наиболее далеко по Охотскому побережью, где по восточным склонам хр. Джугджур достоверно прослежен до 59° с. ш. (Снигиревский, 1946). Севернее они, видимо, встречаются реже, но все же доходят до Охотска, где, по данным Розова (1938), дикуш нет, но, по сообщениям бывших работников золотых приисков, которыми я располагаю, она была нередкой в 1932-1936 гг. по р. Кухтуй (Потапов, 1970а). От Охотского побережья граница, пересекая хр. Джугджур, идет на запад, к пос. Нелькан на р. Мае (кол. ЗИН АН СССР) и далее к среднему течению р. Алдан. По данным О. В. Егорова (устное сообщ.), дикуша встречается по всем аянским ельникам вдоль этой реки, начиная от устья р. Тимптон и вниз по крайней мере до низовьев р. Маи (добыта в 60 км южнее Усть-Маи; Перфильев, 1975), т. е. почти достигая здесь 60° с. ш. Возможно, что дикуша идет еще дальше на север, поскольку существует указание Миддендорфа (1851) на встречи этой птицы местными охотниками где-то по дороге из Якутска в Охотск. По указанию того же автора, наиболее западная точка встречи дикуши — р. Шилка. В качестве западной границы ареала указываются также верховья р. Горбицы, 119° в. д. (Radde, 1863; расспросные сведения), однако самым западным достоверным местом нахождения являются верховья р. Нюкжи (Тугаринов, устное сообщ.) на северном склоне Станового хребта и истоки р. Безымянной — левого притока верхнего Ольдоя — на южном склоне (Гассовский, 1927), т. е. по Становому хребту дикуша идет на запад до 122° 30' в. д. Южная граница ареала проходит по южным склонам хребтов Тукурингра и Станового. Она захватывает верховья р. Уркан, пересекает Зею у устья Гилюя, верховья рек Нора и Депа, р. Селемджу у Селемджинска и верховье р. Бысса (Юдаков, 1972). Далее на восток граница охватывает бассейн р. Бурей, Буреинский хребет, бассейн р. Кур, затем под 50° с. ш. переходит на правый берег Амура и отсюда ареал вдается глубоким клином на юг, по таежному поясу Сихотэ-Алиня, до верховьев рек Большая Уссурка и Рудная, а может быть, и до верховьев Партизанской (Шульпин, 1936). Во всяком случае дикуша вполне обычна в верховьях р. Арму, правого притока р. Большая Уссурка (Михайловский, 1972а). По восточным склонам Сихотэ-Алиня граница идет на север, выходя в районе оз. Кизи на побережье Татарского пролива. Распространение дикуши на Сахалине изучено плохо. Она населяет остров от самых северных его частей примерно до широты г. Долинска (47°30 с. ш.). В связи с повсеместной вырубкой темнохвойных лесов на острове всюду стала очень редкой. На Шантарских островах дикуши нет, несмотря на наличие здесь подходящей растительности.
Климатические условия на протяжении ареала далеко неоднородны по причине не только широтной, но и высотной зональности. Средняя температура января колеблется в пределах от -16 до -32 °С, июля — от 12 до 16 °С. Сумма годовых осадков составляет 500-1000 мм, большинство которых выпадает в летнее время. Снежный покров держится 170-220 дней, высота его в среднем 50-70 см. Длительность безморозного периода 75-105 дней.
Места обитания.
Предпочитается темнохвойная тайга охотского типа, где аянская ель и белокорая пихта (Abies nephrolepis) либо преобладают, либо вкраплены отдельными пятнами среди сплошных лиственничных лесов. Наиболее типичное место обитания — сильно захламленная темнохвойная тайга, на склонах гор или сопок с покровом из мхов или багульника (Ledum palustre). В бассейне нижнего Амура дикуши держатся весной обычно в ельниках у вершин сопок, где ель и пихта растут вперемежку с кедровым стлаником и березой, а кедровый стланик образует местами непролазные заросли. Здесь же птицы и гнездятся. Приверженность дикуши к резко расчлененному ландшафту, где нередки выходы скал и осыпи, прослеживается практически по всему ареалу и отражена в местных названиях вида — каменушка, каменный рябчик. Подрастающие выводки выходят на брусничники и могут далеко уходить от ельников, обитая в чистых лиственничниках и даже на марях. Поздней осенью дикуши уходят в темнохвойную тайгу, где и проводят всю зиму, держась на участках леса на подветренных склонах или в глубоких распадках. На Становом хребте в верховьях Зеи дикуши также живут в верхнем поясе тайги и в поясе кедрового стланика, а в бассейне Селемджи — в верховьях ключей и на северных склонах водоразделов вверх до гольцовой зоны (Шульпин, 1936; Юдаков, 1967; Воронов, 1977). В верховьях Большой Уссурки обычные места обитания — густые ельники, а у бухты Таба летом — и чистые лиственничники с густым подлеском из багульника и моховым покровом (Шульпин, 1936). В верховьях р. Арму основные места обитания — пихотово-еловые леса по водоразделам ключей (Михайловский, 1972а). В верховьях р. Бикин дикуши весной и летом живут в сырых холодных распадках, густо заросших темнохвойной тайгой, где 80% почвы скрыто моховым покровом (А. С. Никаноров, устное сообщ.). В верхнем Приамурье, по р. Горин, так же как и на Сахалине, дикуши населяют смешанные леса из лиственницы и ели как с моховым покровом, так и без него (Мишин, 1959; Абрамов, 1962). На Сахалине, например, в районе с. Дружба в сентябре пара дикуш была встречена в типичных тундроподобных ландшафтах с зарослями кедрового стланика и редкостойными угнетенными лиственницами (Воронов и др., 1975). В центральном Сихотэ-Алине в мае дикуши держались на брусничниках в гольцовой зоне, где прилежащие склоны были покрыты в основном лиственничниками со старыми гарями. Попадались эти птицы и в криволесье из каменной березы, березового и кедрового стланика, в зарослях рододендрона и ряда горных растений на высотах порядка 1300-1500 м над ур. м., где всюду обильны были брусничники с прошлогодней ягодой. В одном случае дикуши были найдены даже в березово-дубовом лесу, но опять же с брусничником по северной стороне гребня. В июле-сентябре дикуши обитали в горных мшистых ельниках до высоты 1600 м над ур. м. и в высокогорных ельниках близ участков мохово-лишайниковой тундры с кедровым стлаником, рододендроном, голубикой, брусникой, а также в елово-лиственничных лесах и на гарях с брусникой (Капланов, 1938). В долине р. Алдан О. В. Егоров встречал дикуш чаще всего на заросших аянской елью сопках в пол склона и очень редко — в хороших ельниках вдоль русел ручьев. Наконец, Юдаков (1972) однажды встретил токующего самца 1 мая в березово-осиновом лесу с подлеском из ольхи в 1 км от ближайшего елово-пихтового леса. Таким образом, места обитания дикуш в летнее время могут быть очень разнообразны, причем приверженность к горам заметно увеличивается по мере движения к югу. Зимой же все птицы концентрируются в елово-пихтовых лесах, дающих им главный зимний корм. Можно поэтому с уверенностью утверждать, что положение северной, а отчасти и западной границ ареала обусловлено наличием достаточно крупных массивов ельников.
Численность.
В силу крайне спорадичного распространения и благодаря очень скрытному образу жизни дикуши в большинстве случаев производят впечатление крайне редкой птицы, нигде не образующей больших скоплений. Все же в целом ряде областей (Алдано-Учурский бассейн, верховья р. Амгунь, бассейн р. Горин, районы оз. Кизи и зал. Де-Кастри, ряд мест на северном Сихотэ-Алине) дикуша является вполне обычной птицей, хотя заметно уступает в численности рябчику. Главная причина низкой численности дикуши — преследование со стороны человека — вблизи вновь возникающих в тайге поселков дикуша сравнительно быстро выбивается начисто. В то же время от таких хищников, как соболь, дикуша в отличие от рябчика терпит очень незначительный урон. Например, в 90 желудках соболей остатки дикуши встречены лишь дважды в начале марта, а в 220 экскрементах этого хищника — ни разу (Юдаков, 1967). Численность в бассейне Селемджи, по данным того же автора, — 2.5 на 1000 га, в верховьях р. Бикин весной — 3 пары на 10 000 га (Никаноров, устное сообщ.). О популяционном составе точных данных нет, но судя по моим наблюдениям, в каждой популяции в весенне-летнее время имеются негнездящиеся годовалые самцы в количестве, несколько превышающем или равном числу взрослых самцов. Эти молодые птицы держатся крайне незаметно и с трудом обнаруживаются.
Территориальность.
Территориальное поведение у азиатской дикуши примерно такого же характера, как у американских видов рода, у воротничкового рябчика и у дымчатых тетеревов. Весной самец занимает определенный участок, где токует и спаривается с приходящими сюда самками. Эти токовые участки расположены в тайге крайне неравномерно, отдельными скоплениями по 2-4, причем некоторые исследователи считают такие скопления аналогами диффузным токовищам обыкновенного глухаря (Пукинский, Никаноров, 1974). Об исключительной оседлости дикуш говорят все более или менее детальные наблюдения. Так, отдельные неразмножающиеся птицы за лето перемещались в пределах буквально нескольких десятков квадратных метров (Никаноров, устное сообщ.). Тем не менее какие-то сезонные перемещения, связанные со сменой характера питания, явно существуют, но они еще не исследованы.
Размножение и токование.
Сроки начала токования неизвестны. В верховьях р. Горин токовавших самцов наблюдали с конца апреля (Абрамов, 1962), a Л. Г. Капланов в Сихотэ-Алиньском заповеднике — 17 мая. Мне пришлось наблюдать токовавшего самца на Нижнем Амуре 11 мая. Видимо, токовой период в южной половине ареала захватывает конец апреля и большую часть мая, на севере ареала сроки несколько более поздние. По наблюдениям А. С. Никанорова (устное сообщ.), токование в верховьях Бикина протекало наиболее интенсивно в первой половине мая, но песни самцов можно было провоцировать до начала июня.
Ток у дикуш, как правило, носит одиночный характер, но в отдельных случаях, когда плотность популяции достаточно велика, самцы токуют в непосредственной близости один от другого, образуя что-то похожее на групповое токовище. К. Г. Абрамов, наблюдая 7 мая токовавшего самца, вспугнул в 20 м от него еще 2 самцов и еще в 10 м — самца и самку. Вполне возможно, что часть этих самцов была молодыми, которые не принимают активного участия в токовании. Например, в районе наблюдений А. С. Никанорова наряду с токующими взрослыми самцами находились и молодые, не принимавшие участия в размножении. Б. Н. Вепринцев (устное сообщ.) наблюдал «в 700 км севернее Хабаровска» групповое токование дикуш, когда в конце апреля несколько самцов токовали в пределах непосредственной видимости и слышимости один от другого (около 100 м); но он не заметил, чтобы дикуши, токовавшие в непосредственной близости друг от друга, провоцировались бы взаимно токовыми ритуалами или вступали в конфликты, т. е. основные элементы группового тока здесь отсутствовали. И, наконец, А. С. Никаноров проводил наблюдения на участке, где токовали 3 самца, причем расстояния между центрами токовых участков были от 150 до 200 м, т. е. эти самцы не видели и не слышали друг друга в процессе токования. Естественно, что при такой ситуации говорить о групповом токе не приходится. Остальные же авторы, дававшие описание токования, вообще наблюдали, только одиночных самцов (Миддендорф, 1851; Капланов, 1938; Yamashina, 1939; Снигиревский, 1946; Потапов, 1969а).
Токовой участок располагается обычно в верхней части сопки или оврага в темнохвойном лесу, часто с моховым покровом. В такой ситуации на более или менее свободном от бурелома месте располагается центр участка, занимающий обычно несколько квадратных метров. Это место обычно изрядно вытаптывается токующим самцом, который выполняет свой ритуал преимущественно здесь, не удаляясь далее 25-30 м. Однако Л. Г. Капланов наблюдал токование на горелом гольце, заросшем багульником (в багульнике токуют и канадские дикуши), березками Миддендорфа и красной смородиной. Вполне очевидно, что в данной ситуации дикуши оказались вторично, после того, как сгорел хвойный лес. Нередко местом токования служит большой пень или колода (Yamashina, 1939), как и у воротничкового рябчика.
Основная токовая поза самца — типичная для большинства тетеревиных птиц (рис. 6). Шея поднята, но голова в нормальном положении — птица смотрит прямо перед собой. Шея выглядит сильно расширенной из-за взъерошенных перьев. Хвост поднят вверх под углом около 80° и слегка расправлен, крылья отставлены в стороны и приспущены, но по земле не волочатся. Расхаживающая в такой позе птица то и дело останавливается и прислушивается. Когда мне пришлось пробираться сквозь глухие заросли к токовавшему самцу, последний начинал активно токовать всякий раз, когда под ногами трещала ветка или вообще издавался шум. Перед выполнением основного токового цикла, который довольно однообразен, самец поднимает максимально раскрытый хвост, взъерошивает перья на затылке, щеках и подбородке и начинает двигаться семенящим шагом, складывая и раскрывая с характерным громким шорохом крайние рулевые перья. А. С. Никаноров отмечал на этой стадии своеобразные движения зоба, создающие впечатление тяжелого дыхания птицы; одновременно с этим начинают вздрагивать хвост и крылья, причем хвост при вздрагиваниях смещается к голове, а крылья в вертикальной плоскости. Движения хвоста и крыльев происходят согласованно, с небольшими интервалами. Шаги самца убыстряются, в такт с шагами несколько раз с громким шорохом распускается полностью и частично складывается хвост (рис. 6). Затем с шорохом раскрыв хвост, птица внезапно останавливается, замирая в основной токовой позе. Начинается мелкая вибрация крыльев, перья на крестце и шее взъерошиваются и вслед за этим самец издает своеобразный вибрирующий звук, вроде «урррррррррр», быстро повышающийся к концу. Звук этот, длящийся около 3 с, несколько напоминает завывание ветра в трубе и слышен на очень небольшом расстоянии. Даже поблизости от токующего самца кажется, что он доносится издалека, и трудно определить, откуда он исходит. Сам звук начинается на низкой гудящей ноте и тянется затем с повышением тона до перехода в отдельные отрывистые звуки, которые, быстро следуя один за другим, и создают эффект вибрации. Тон этих отдельных звуков, число которых 39-40, также продолжает повышаться вплоть до последнего, тональность которого на 4 ноты выше первого звука (рис. 7). Длительность первого гудящего звука примерно 1 с, а остальных, следующих за ним, не превышает 0.05 с, при интервалах 0.01 с — человеческим ухом они воспринимаются как один вибрирующий звук. Голову при этом самец держит прямо, клюв закрыт, крылья совершенно неподвижны. Последнее обстоятельство доказывает исключительно вокальный характер звука, в то время как Снигиревский (1946) предполагал, что звук этот происходит от быстро вибрирующих крыльев самца. Закончив урчание, птица почти сразу совершает два коротких, не свыше 30 см, прыжка, хлопая при этом крыльями, бьющими по воздуху и по земле, и щелкая наподобие каменного глухаря.
Замедленное прослушивание записи токования и анализ ее сонограммы, любезно присланной мне доктором Бергманном (рис. 7), убедили меня в том, что эти щелчки продуцируются не голосовым аппаратом, как у глухарей, а крыльями — щелчки и взмахи крыльев синхронны. Совершенно такие же щелчки, но одиночные, издает горная дикуша во время токового полета: там было точно установлено, что эти щелчки — результат столкновения крыльев над спиной птицы при особенно глубоких взмахах (см. далее). Такое примечательное сходство в токовании этих двух видов безусловно не случайно и говорит об их близком родстве.
По наблюдениям А. С. Никанорова, во время первого прыжка птица делает разворот на 180°, смещаясь в сторону, а второй прыжок выполняется на месте (Пукинский, Никаноров, 1975). Первый щелчок слышен в момент первого прыжка и иногда сдваивается (рис. 6), а при втором прыжке издаются два двойных щелчка. Все это звучит примерно, как «урррр... так... так-так... так-так». Щелчки — самые громкие звуки тока и слышны на расстоянии до 100 м. Урчание же едва слышно уже в 30-40 м от птицы. Токующий самец, которого я наблюдал в низовьях Амура, после второго прыжка падал на грудь, замирая на секунду в такой позе с частично распластанными крыльями и вертикально поднятым и даже слегка запрокинутым на спину хвостом. У горной дикуши есть аналогичная поза «присаживания» (Hjorth, 1970). Закончив ритуал этой позой, самец привставал, подбирал крылья, взъерошенное оперение опускалось и птица возобновляла кружение по площадке. Этот ритуал повторяется с различными интервалами, минимальный — 18 с.
Токовые полеты у азиатской дикуши есть, но выполняются редко. По наблюдениям А. С. Никанорова, самец перед токовым полетом принимает позу угрозы и издает негромкий звук «хр-хр», после чего взлетает вверх на высоту 2-3 м трепещущим полетом с зависанием в верхней точке полета. Спуск происходит так же, как и подъем, а при приземлении слышно несколько щелчков. Временами этот полет может усложняться, как у американских дикуш, — самец взлетает на ветку дерева, вновь слетает вниз, продолжая токование на земле. Л. Г. Капланов, наблюдавший этот взлет на дерево, пишет, что самец там принимал позы «наподобие токующего глухаря» и «с громким шорохом распускал хвост». Сообщение о том, что самец может петь на дереве (Перфильев, 1975) явно ошибочно. Пение и прыжки со щелчками являются неразделимыми частями одного ритуала, выполнение которого на сучке невозможно. А. С. Никаноров также пишет, что самец петь на дереве не может.
Имеются вполне определенные указания на то, что азиатская дикуша — полигам (Пукинский, Никаноров, 1974). По наблюдениям А. С. Никанорова, самка посещает самца на токовом участке по своей инициативе и, когда она его покидает, тот не уходит за ней, но с новой энергией возобновляет токование. Токование протекает наиболее активно утром, на восходе солнца.
Данных об осеннем токовании мало. Л. Г. Капланов наблюдал токовавшего на земле самца в хвойном лесу 31 августа.
Гнездование.
Данные о гнездовании азиатской дикуши в последнее время значительно пополнились благодаря наблюдениям А. Г. Юдакова и А. С. Никанорова.
Юдаков (1972) приводит данные, сообщенные ему охотником В. Ф. Гребневым о гнездовании дикуш, которое последний наблюдал в течение двух лет у своего зимовья. «Им было найдено гнездо дикуши под валежиной в елово-пихтовом лесу в долине небольшого ключа. Лоток был выстлан кусочками зеленого мха, травой и листьями. В день нахождения гнезда в нем было 1 яйцо, через 6 дней (наблюдения проводились раз в неделю) — 6 яиц, т. е. откладка яиц происходила ежедневно. При последующих посещениях самка сидела на гнезде и не слетала даже тогда, когда к ней прикасались рукой. Через 5 недель после первого посещения около гнезда был встречен выводок из 12 птенцов. Болтунов в гнезде не оказалось, т. е. полная кладка состояла минимум из 12 яиц. Во время посещения гнезда самец неподвижно сидел поблизости на дереве, а позднее он всегда держался вместе с выводком. Самка не отводила человека от выводка, а затаивалась или медленно передвигалась по земле. При тревоге птенцы разбегались и затаивались. При этом их легко можно было взять руками, а самка проявляла лишь едва заметную тревогу, медленно продвигаясь в сторону человека с тихим звуком, напоминающим квохтание курицы. Даже перепархивающих с дерева на дерево птенцов можно было легко поймать. Во время кормежки птенцы медленно передвигались по земле, время от времени что-то склевывая с листьев и ловя в воздухе насекомых, но в земле не рылись. Выводок находился не далее 50-100 м от гнезда. Всю зиму выводок держался на гнездовом участке. Весной стайка разбилась на пары. На этом участке было отмечено 4 пары дикуш. 1 июля было найдено гнездо в основании наклонного дерева. Кладка состояла из 11 яиц. На следующий день в гнезде оказалась только скорлупа, а поблизости был обнаружен весь выводок, т. е. вылупление птенцов произошло в течение 1 суток. Выводки не более недельного возраста встречены 4, 7 и 10 июля 1967 года. Последний состоял из 12 птенцов.
Очень интересные данные сообщает А. С. Никаноров, нашедший гнездо в разреженном ветровалом лесу, где почву покрывал толстый слой мха. Гнездо представляло собой ямку, вытоптанную во мху с юго-восточной стороны ствола дерева, диаметром 170 и глубиной 70 мм. Оно было выстлано сухой кедровой хвоей, толщина выстилки — 15 мм. Под выстилкой мох был снят до самой почвы.
Гнездо было устроено в сыром месте, хотя вокруг было порядочно вполне сухих участков. В гнезде находилось 7 яиц, их окраска — бледно-охристый тон с каштановыми крапинками и пятнами, более крупными на тупом конце: средний диаметр таких пятен — 2-3 мм, максимально до 5 мм. Масса яиц, уже насиженных, колебалась в пределах 22-23.5 г, их размеры: 44x33, 45x33, 46x36, 46X36, 48X36, 46X36, 48X34 мм. Самка насиживала очень плотно, позволяя брать себя руками, однако при этом однажды выскочила из рук наблюдателя, оставив в его руках значительную часть своих перьев. Самка на гнезде при приближении опасности издавала рулевыми перьями шорох, как и токующий самец, а кроме того испускала особый завывающий звук, напоминающий «песню» самца — «у-уууу… у-ууу... у-уииииии...». Очень важно отметить, что, когда гнездо это оказывалось на солнце (между 9 и 12 ч), птица явно страдала от жары.
Масса вылупившегося птенца 16-17 г. Птенцы вылупляются с 7 трубками первостепенных маховых перьев и 10 второстепенных. Радужина у них темно-бурая, клюв и ноги — светло-стальные, пенек 8-го первостепенного махового появляется на 8-9-й день. Голос птенцов — высокий призывный писк, как и у остальных тетеревиных. В поисках насекомых они осматривали листья с нижней стороны и клевали снизу вверх (аналогичное наблюдение Н. Н. Данилова за птенцами рябчика). Ритмика дневной активности состояла из чередования коротких кормежек (5-10 мин) и отогревания под самкой (10-15 мин). Птенцы ночевали под матерью до месячного возраста. В 4-дневном возрасте они уже подпрыгивали, трепеща крыльями, а в 7-8-дневном уже могли взлетать на нижние ветки деревьев. В возрасте с 3 до 10 дней птенцы были очень пугливы, затаивались, затем же вновь стали доверчивыми. Самка при беспокойстве издает негромкое квохтание, созывает же птенцов негромким «квох... квох...». Самцов вблизи гнезд или выводков А. С. Никаноров не наблюдал. Возможно, что в случае, описанном А. Г. Юдаковым, вблизи самки находился молодой самец, не принимавший участия в размножении.
Единственное гнездо с о-ва Сахалин, подробное описание которого дано в литературе, было найдено на южном Сахалине. Оно располагалось в густом, слегка заболоченном хвойном лесу из сахалинской пихты и аянской ели с примесью даурской лиственницы. Гнездо помещалось на свободном от мха участке земли и было скрыто в зарослях Vaccinium chamissonis и V. smallii. Гнездо это представляло собой ямку, небрежно выстланную веточками пихты и листьями Vaccinium spp. 8 июня оно содержало 8 яиц. Примерная степень насиженности — 3 дня. Цвет яиц палево-коричневый, с легким оливковым оттенком и разбросанными всюду коричневыми пятнышками. Размеры их в мм: 48X32, 45.5x32.46, 46x32, 45x32, 46x32, 48x31.5, 46x32, 43x31. Второе гнездо было найдено у подножья пихты на поляне, где деревья росли разреженно (Taka-Tsukasa, 1932).
Спаривание на центральном Сахалине наблюдалось во второй половине мая (Гизенко, 1955). 16 мая в верховьях р. Горин добыта самка с готовым к сносу яйцом (Абрамов, 1962). Для бассейна Селемджи начало кладки указано в конце мая-начале июня (Юдаков, 1967). Видимо, кладка происходит во второй половине мая, и в конце этого месяца самки садятся насиживать. В упомянутом выше гнезде, найденном на Сахалине, самка приступила к насиживанию примерно 5 июня. В конце июня появляются птенцы. Маленьких птенцов можно встретить и в конце июля (Афанасьев, 1934). Данных о длительности насиживания нет. А. С. Никаноров отметил вылупление птенцов в 3 гнездах 16, 20, 21 июня. Наиболее ранняя дата встречи птенцов дикуши на Сахалине — 22 июня (Yamashina, 1939); 25 июня самка с пуховым птенцом встречена в верховьях Та-Кунжи, в Сихотэ-Алиньском заповеднике (Капланов, 1938), 27 июня выводок из 5 пуховых птенцов был встречен в верховьях р. Горин (Абрамов, 1962). Там же 7 июля замечен выводок с птенцами в ювенальном оперении; размеры одного из этих птенцов — крыло 120 мм, хвост 40 мм. 31 июля молодые самец и самка, добытые из выводка в 5 штук, имели массу соответственно 250 и 248 г, размер крыла — 140 и 160 мм, длину хвоста — 55 и 40 мм. Шульпин у бухты Таба 4 августа встретил выводок из 8 птенцов, размером уже в серую куропатку. Материалы ЗИН АН СССР: 23 июля, Лантарь (Охотское побережье), крыло 120 мм, полный ювенальный наряд, начали расти ПМП-9 и ПМП-10, отрастает дефинитивное ПМП-2, немного пуха сохранилось только на горле. 26 июля, Туксаин (Становой хребет) — моложе предыдущего, крыло 103 мм, в наличие только 8 маховых перьев, из которых первое — дефинитивное — только начинает отрастать, на голове еще пуховой наряд. 31 июля, верховья р. Амгунь, марь: крыло 125 мм, полный ювенальный наряд, выпало ювенальное ПМП-3. 5 августа зал. Де-Кастри, 3 птенца; у двух самок (крыло 155 и 156 мм) растут ПМП-9 и ПМП-10, первые 6 ПМП уже сменились на дефинитивные, хвост у одной еще ювенальный, у другой отрастают дефинитивные рулевые, у третьего птенца крыло 145 мм, ПМП-10 еще в пеньке, сменились на дефинитивные только 5 ПМП. 2 птенца от 13 августа со Станового хребта: крылья соответственно 153 и 157 мм, сменились ПМП-6 и ПМП-7, контурные перья дефинитивного наряда появились на боках тела и груди. И, наконец, молодая самка от 14 сентября (Немуй, Охотское побережье) заканчивает линьку в первоосенний наряд: еще не доросло ПМП-8, остались ювенальными плечевые перья, длина хвоста 96 мм. Пуховой птенец дикуши, хранящийся в Зоомузее МГУ, добыт 15 июля у берега Ульбинского залива, в 150 км от Николаевска.
Эти отрывочные данные позволяют заключить, что массовое вылупление птенцов происходит в конце июня-начале июля и к 80-дневному возрасту молодые птицы практически достигают размеров взрослых и заканчивают линьку в первоосенний наряд. Птенцов водит обычно самка, иногда вместе с самцом. Выводок очень длительное время держится на одном и том же месте, на ограниченном пространстве радиусом не более 100 м от гнезда (Taka-Tsukasa, 1932; Юдаков, 1967). Японские авторы указывают на высокую способность пуховых птенцов дикуши к полету: птенцы с едва сформировавшимися крыльями взлетают на макушки деревьев на высоту до 20 м над землей. Встреченные К. Г. Абрамовым 27 июня пуховыё птенцы вспорхнули и с писком расселись на лиственнице (как указывал еще Шульпин, птенцы дикуши пищат, как тетеревята). О возрасте этих птенцов ничего не сказано, но фотография пятидневных птенцов в работе Ямасины (Yamashina, 1939) показывает, что крылья у них в этом возрасте вполне пригодны для полета. И, наконец, А. С. Никаноров вполне определенно говорит о том, что птенцы способны взлетать на деревья с 7-дневного возраста.
Хозяйственное значение и охрана.
Азиатская дикуша внесена в «Красную книгу СССР» как вид, численность которого быстро сокращается и которому грозит полное истребление по причине описанных выше особенностей его поведения. Даже в прежние времена этот вид не имел никакого промыслового значения. Из-за небольшой плотности населения биоценотическая роль дикуши также явно невелика и как консумента, и как продуцента. Однако именно особенности ее поведения делают дикушу очень перспективным объектом для разведения в ландшафтах, заметно измененных антропогенным воздействием, в окультуренных, лесопаркового характера участках тайги, которых в пределах ареала становится все больше.
Проблемы охраны в настоящее время сводятся исключительно к организации осуществления действенного запрета на добычу этих птиц, на усиление пропаганды охраны вида. Однако уже назрело время для срочной разработки методики содержания и размножения в неволе для последующего расселения, задачи сложной, но вполне выполнимой. Успешные опыты по разведению в неволе американских дикуш уже проведены (Pendergast, Boag, 1971а).
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные.", 1985 г.
 Дикуши. Род Falcipennis.
Дикуши. Род Falcipennis. Канадская дикуша. Falcipennis (canachites) canadensis.
Канадская дикуша. Falcipennis (canachites) canadensis.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
.") Обыкновенный рябчик (Bonasa bonasia).
Обыкновенный рябчик (Bonasa bonasia). sewerzowi.") Рябчик Северцова. Bonasa (Tetrastes) sewerzowi.
Рябчик Северцова. Bonasa (Tetrastes) sewerzowi. Воротничковый рябчик. Bonasa umbellus.
Воротничковый рябчик. Bonasa umbellus. Рябчики.
Рябчики. Суточный бюджет энергии и времени тетеревиных птиц.
Суточный бюджет энергии и времени тетеревиных птиц.
