Канадская дикуша. Falcipennis (canachites) canadensis.
Лесная тетеревиная птица несколько крупнее рябчика, с незначительным половым диморфизмом в окраске и размерах.
Образ жизни.
Питание.
Зимний сезон в жизни канадских дикуш.
Размножение и токование.
Гнездование.
Окраска и оперение.
Размеры и масса.
Распространение и ареал.
Места обитания.
Численность.
Популяционный состав.
Территориальность.
Географическая изменчивость.

Канадская дикуша (самка).
Образ жизни.

Дикуша канадская (самец).
Наиболее яркая черта в поведении канадской дикуши, как и у остальных дикуш, — отсутствие боязни человека. Так же, как и азиатская дикуша, канадская может подпустить человека почти вплотную и дать накинуть на себя петлю. Это тихая, малозаметная птица, ведущая преимущественно наземный образ жизни и перемещающаяся большей частью пешком. Полет такой же шумный и маневренный, как и у рябчиков. В случае опасности птицы предпочитают уходить пешком и взлетают с явной неохотой. Незаметности птиц способствует их молчаливость. Вне брачного сезона для самца вообще неизвестно никаких голосовых реакций, за исключением разве что тонких клохчущих звуков (Bent, 1932). Для самки описан громкий визгливый позыв «киррр» при тревоге (Johnsgard, 1973), а также мягкие клохчущие звуки, издаваемые обеспокоенной самкой на дереве или же при выводке, звучащие как «крук... крук... крук», среди которых и проскальзывает более раскатистое «кррррук» (Bent, 1932).

Рис. 1. Канадская дикуша Falcipennis canadensis, самец. 1 — вид со спины; 2 — с брюшной стороны.
Питание.

Рис. 2. Перо с грудки Falcipennis canadensis.
Зимнее питание по всему обширному ареалу базируется почти исключительно на иглах елей и сосен. По всей Аляске (Фербенкс, п-ов Кенай, оз. Алекнагик) основу зимнего питания составляет хвоя черной и белой ели с октября по конец апреля включительно (Ellison, 1966). В центральной Альберте, Канада, главный зимний корм — хвоя сосны Pinus contorta, которая преобладает в корме и весной (Pendergast, Boag, 1970). На востоке ареала (Онтарио, Канада и Мичиган, США) главным зимним кормом является хвоя сосны Бангса (Crichton, 1963; Gurchinoff, Robinson, 1972). Весной по мере стаивания снега в корме начинают преобладать прошлогодние ягоды брусники и вороники, листья и цветки различных растений (в том числе листья голубики и березы), лишайники. В центральной Альберте птицы поедают весной помимо сосновой хвои также и еловую (встречаемость в зобах до 29%). В летней диете важное место занимают верхушечные побеги и стебли хвоща, семена осок, свежерастущие концевые пучки еловой хвои, но преобладающим кормом являются листья и в меньшей степени цветки и ягоды Vaccinium membranaceum. Летом по-прежнему употребляется много старых ягод, а также насекомых и пауков. В осеннем рационе наряду с большим количеством ягод с сентября быстро увеличивается доля хвои, которая уже в ноябре составляет 92.5% рациона. На востоке ареала в октябре большую роль в питании играет хвоя лиственницы — вплоть до ее опадания (Crichton, 1963; Pendergast, Boag, 1970, 1971b,). Напомним, что такой корм в это же время характерен и для обоих видов глухарей. Переход на хвойную диету осенью совершается часто при отсутствии снежного покрова (Keppie, 1977).
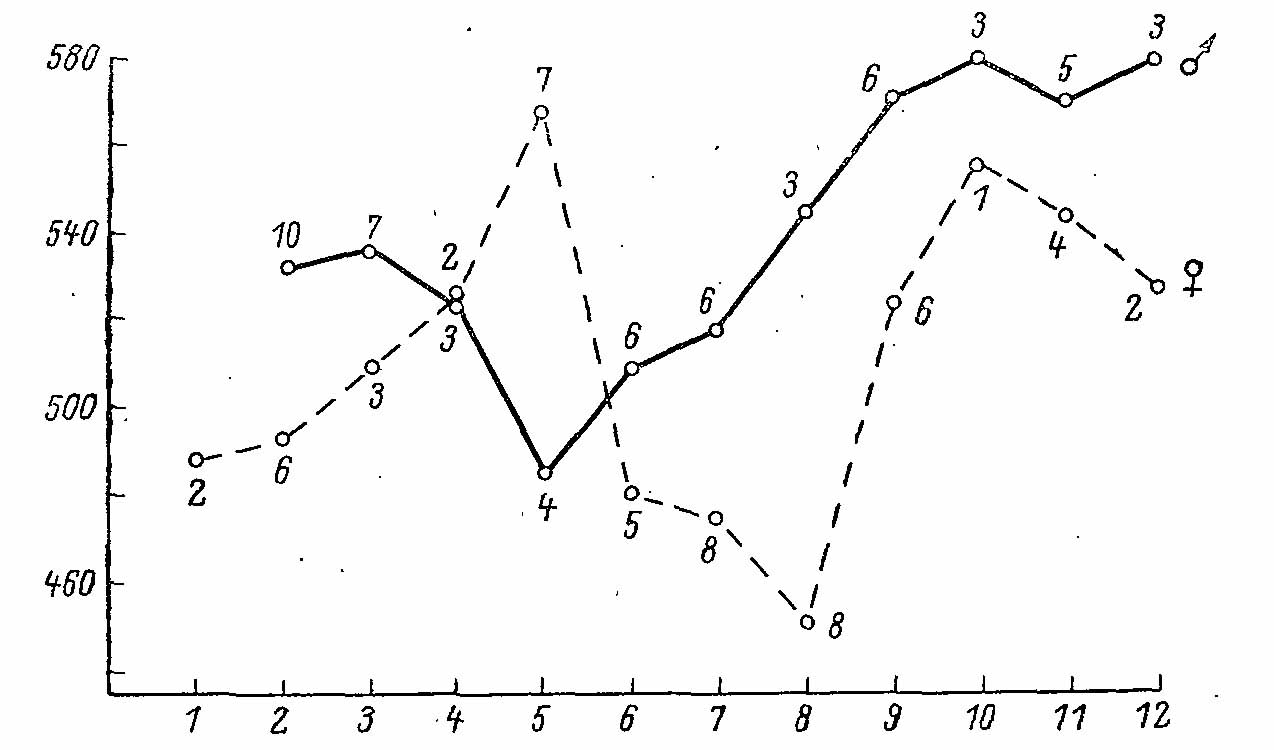
Рже. 3. Сезонная динамика массы Falcipennis canadensis. По оси абсцисс — месяцы; по оси ординат — масса, в г. Цифры — число экземпляров (по Pendergast, Boag, 1973).
Зимний сезон в жизни канадских дикуш.
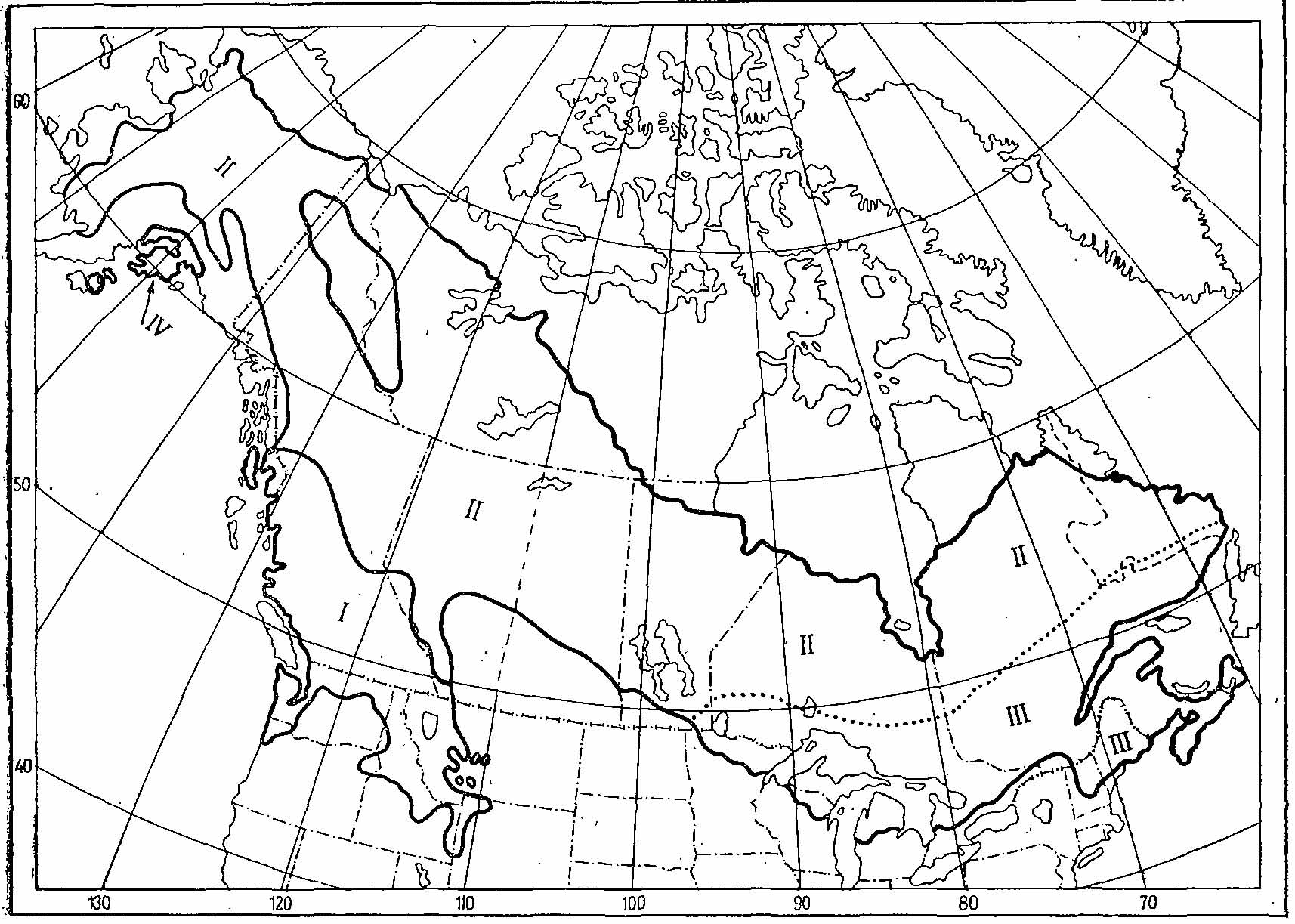
Рис. 4. Ареалы Falcipennis canadensis и F. franklinii (по Aldrich, 1963; Johnsgard, 1973. I — F. franklinii; II — F. canadensis canadensis; III — F. canadensis сanace; IV — F. canadensis atratus.
О зимнем образе жизни сведений практически нет. В то же время зимний бюджет энергии был выяснен недавно полевыми наблюдениями в сочетании с изучением птиц, содержащихся в вольерах, в центральной Альберте. Величина суточного рациона, состоящего из сосновых игл, составила 40.4 г сухой массы или же, принимая содержание воды в иглах за 50%, 80.8 г сырой хвои (899.8 кДж/сутки). Из этого количества птицы усваивали только 10.9 г сухого вещества. Величина экскреторной энергии в течение суток — 623.0 кДж/сутки, т. е. энергия существования составляет 276.8 кДж/ сутки. Столь низкая величина энергии существования объясняется, видимо, тем, что она исследовалась на вольерных птицах, подвижность которых была резко ограничена. Разница в энергии существования свободноживущих и вольерных тетеревиных птиц, как показали работы на белой куропатке, составляет 20.8% от энергии существования последних (West, 1968). Эта «стоимость свободной жизни» должна быть примерно равной у тетеревиных птиц одной весовой категории и в таком случае энергия существования канадской дикуши будет равна 334.4 кДж/сутки, что все же существенно ниже, чем у равной ей по массе белой куропатки (489.9 кДж/сутки; West, 1968). Возможно, что основной выигрыш, получаемый дикушами от медлительного, мало подвижного образа жизни, заключается именно в этом.
Размножение и токование.
Сроки размножения поздние. Наиболее активное токование самцов происходит в первой половине мая, в середине мая самки заканчивают кладку и приступают к насиживанию, в конце первой-начале второй декад июня появляются птенцы. В Нью-Брунсвике, Канада, период вылупления в зависимости от хода весны может быть между 10 и 28 июня или между 14 июня-1 июля (Quinn, Keppie, 1981). На северной окраине ареала сроки размножения запаздывают почти на месяц — на п-ове Унгава, к примеру, откладка яиц начинается около 5 июня, а насиживание — около 12 июня (Bent, 1932). Самцы токуют на своих токовых участках, расположенных обычно в достаточно густом лесу, но с наличием участков с разреженным древостоем, где регулярно выполняют токовой ритуал и спариваются с появляющимися вблизи самками.
Токовой ритуал состоит из токового полета и ряда дисплеев, проделываемых на земле. В основной токовой позе самец стоит с поднятой головой, слегка выпятив грудь и подняв слегка раскрытый хвост под углом 70°. Нижние кроющие перья хвоста также подняты, и их белые вершины образуют характерный узор на чёрном фоне рулевых перьев. «Брови» расширены и возвышаются над головой двумя валиками. Перья на шее (кроме задней поверхности) и передней части груди подняты, благодаря чему птица кажется более широкой, а белые концы перьев на груди образуют белую поперечную перевязь на черном фоне. Находясь вблизи самки, а временами и в одиночку, в основной токовой позе, самец выполняет довольно сложный ритуал, во время которого он делает небольшую пробежку из ряда ускоряющихся шагов. Останавливаясь, он продолжает топтаться на месте и с наклоном головы издает короткое резкое шипение и сразу же за ним — высокий визгливый звук. Опуская голову, самец взъерошивает перья также и на задней стороне шеи, а с началом шипения хвост раскрывается на 3/4 и тут же складывается и вновь раскрывается уже полностью с громким шорохом. При остановке самец одновременно опускает к земле слегка раскрытые крылья. Другой ритуал, также обычно совершаемый перед самкой, заключается в резких подергиваниях головой то в одну, то в другую сторону, причем с каждым рывком головы хвост раскрывается и складывается с громким шорохом. При этом птица слегка приседает на лапах, приподнимает крестец, слегка раскрывает крылья и топорщит перья на груди.
Токовой полет может совершаться в различной манере, но здесь можно выделить два основных типа. В одном случае токовой полет напоминает скорее токовой взлет — птица поднимается на быстро трепещущих крыльях вертикально вверх, до 5-метровой высоты и таким же образом опускается вниз, делая краткую задержку в верхней точке полета. В других, достигнув верхней точки полета, птица садится на сучок дерева и только после некоторого перерыва, длящегося от нескольких секунд до нескольких минут, слетает на землю. Обоим этим типам полета свойственна одна общая черта — перед опусканием на землю, когда до нее остается 1-2 м, птица резким движением ставит свое тело почти вертикально, широко раскрывает хвост и приземляется на усиленно бьющих крыльях. На земле самец сразу же принимает основную токовую позу. На ветке дерева самец сидит в спокойном состоянии и может здесь даже кормиться. Хлопанье крыльев при посадке бывает очень громким и слышно до 200 м.
При атаке соперника самец вытягивает шею, складывает и опускает хвост, прижимает оперение к телу и максимально расширяет «брови». При беге навстречу сопернику он издает серию коротких и резких шипящих звуков — два в первую половину секунды и остальные пять — за полторы секунды. С каждым из этих звуков хвост также открывается и складывается с отчетливым шорохом (описание токового поведения дано по Lumsden, 1961а, с добавлениями из Hjorth, 1970).
Есть данные, что самцы могут принимать основные токовые позы в течение всего года (Robinson, Maxwell, 1968; Ellison, 1973), особенно осенью в период осеннего токования. Токовые полеты осенью выполняются очень редко.
Гнездование.
Гнездо — обычного для тетеревиных птиц типа, представляет собой небольшую ямку диаметром около 20 см, скудно выстланную сухими листьями, хвоей, травой и небольшим количеством перьев, выпавших у наседки. Нередко гнездо устраивается в толстом слое мха (Bent, 1932). Гнезда располагаются большей частью под прикрытием нависающих еловых ветвей и бывают очень хорошо скрыты (Johnsgard, 1973). Незаметности гнезда способствует и покровительственная окраска самки, и ее поведение — она насиживает очень плотно позволяя даже трогать себя руками. Описан случай, когда вокруг насиживающей самки ходили люди, неоднократно бегали белки, подлетала вплотную кукша — самка не реагировала на это ни одним движением (Walkinshaw, 1948). Для гнездования выбираются как самые захламленные участки леса, так и достаточно разреженные участки с редким подлеском, иногда даже на опушках. Гнезда могут быть расположены на самом различном расстоянии друг от друга, известен случай гнездования двух самок почти рядом друг с другом (Palmer, 1949). Окраска яиц красновато-коричневая, с небольшим количеством разбросанных по всей поверхности резко очерченных пятен и точек, причем крупных пятен, размером с горошину, мало и цвет их темно-коричневый или ржаво-красный (Short, 1967). По наблюдениям в неволе, окраска основного фона яиц дикуши такая же, как у молодых почек ели, которыми охотно питается самка, и чем больше она их ест в период откладывания яиц, тем гуще и сочнее их окраска (Bishop, 1912). Покидая гнездо с неоконченной кладкой, самка каждый раз закрывает ее подручным материалом, чего уже не делается при насиживании (Bent, 1932).
Самка откладывает яйца с интервалом от 1 до 3 суток (наблюдения в неволе; Pendergast, Boag, 1971а). Количество яиц в кладке от 4 до 12, обычно же в пределах 6-9 яиц. На севере кладки более крупные, чем на юге. Например, в Новой Шотландии, Канада, средняя величина кладки 5.8 яиц (по 39 кладкам, с колебаниями от 4 до 10 яиц; Tufts, 1961), а на Аляске — 7.5 (по 26 кладкам, с колебаниями от 4 до 9 яиц; Ellison, 1974). Размеры яиц (107 штук): 43.5x31.1 мм (Bent, 1932).
Срок насиживания, определенный в 2 случаях в природе, оказался равным 23.4 дням (24 дня, по данным Palmer, 1949), на 2.5 суток дольше, чем при вольерном разведении (21 день; Pendergast, Boag, 1971а). Режим инкубации не прослежен. Самка может покидать гнездо, отправляясь на кормежку, взлетая прямо с него, и по возвращении, сев поблизости на дерево и убедившись в отсутствие опасности, приземляться прямо на гнездо (Walkinshow, 1948).
Целый ряд наблюдений показывает, что самец может продолжительное время обитать вблизи гнезда, а также у места жизни выводка, но никакого участия в охране гнезда или выводка не принимает.
Первые две недели жизни птенцы прибавляют в массе по 1.61-2.55 г в день, во 2-5 недели — по 4.82-6.71 г в день и далее до 2-месячного возраста — по 4.31-5.93 г (Quinn, Keppie, 1981). Выводки сравнительно подвижны, и величина участка обитания может варьировать от 6.1 до 155 га. После распада выводков, который происходит в сентябре, на месте гнездования остается только 9% молодых птиц, остальные же разбредаются по окрестностям, но далеко не уходят. Максимальное удаление в первую осень — 10.5 км (Ellison, 1973).
Хозяйственное значение канадской дикуши довольно высоко, в основном в качестве объекта спортивной охоты. По уровню добычи она занимает (вместе с горной дикушей) третье место — 440 000 в осенний охотничий сезон 1970 г. (Johsgard, 1973).
Окраска и оперение.
Взрослый самец.
Общий тон окраски верха черновато-серый, с правильным поперечно-полосатым рисунком (нередко дугообразной формы) из оливково-серых вершинных полос. Эти полосы особенно широки на шее и голове, создавая здесь почти сплошную оливково-серую поверхность. Окраска перьев верхней части тела однообразна — на черноватом фоне в верхней половине пера несколько поперечных полос, из которых верхняя оливково-серого цвета, а остальные — светло-коричневые. На нижней части тела выделяется черное, с редкими белыми пестринами горловое пятно, имеющее нечеткую белую окантовку, начинающуюся, как и у рябчика, позади черных ноздрёй, но прерывающуюся у глаза. За глазом — небольшое белое пятно, имеющее неясное продолжение в виде едва намеченной полоски. Шея спереди оливково-серая (летние перья в июле-августе придают ей коричневый оттенок), с четкими белыми наствольными полосками на перьях, прилегающих к белой окантовке горла. Грудь черная, с узкими белыми вершинными полосами перьев в верхней ее части и широкой белой окантовкой в нижней (рис. 2). Эта окантовка, создаваемая широкими белыми вершинами перьев, отделяет (иногда не полностью) черный грудной пластрон от черного пятна в центре передней части брюха, также окруженного со всех сторон белыми пятнами вершинных полос. Бока сероватые, перья здесь несут клинообразные наствольные вершины белого цвета, особенно крупные на самых длинных перьях, и серый поперечнополосатый рисунок по черновато-бурому фону. Окраска ПМП (первостепенные маховые перья) черно-бурая, с белесыми краями наружных опахал (иногда с коричневыми поперечными полосами) на ПМП-5-ПМП-8 и сероватой вершинной окантовкой на ПМП-1-ПМП-8. ВМП (второстепенные маховые перья) такого же цвета, с более широкой беловатой окантовкой вершины пера и нечеткими коричневатыми поперечными полосами на наружном опахале внутренних перьев. На плечевых перьях — клинообразные вершинные белые пятна и размытый струйчатостью желтовато-серый поперечный рисунок. ВКК (верхние кроющие крыла) несут такой же неясный поперечный рисунок желтовато-серого цвета по черно-бурому фону, изредка на них встречаются узкие, едва заметные стержневые полоски белого цвета. РП (рулевые перья) черные с широкими рыжими пятнами на вершинах, имеющими тонкую, часто неполную черную окантовку. ВКК отличаются от перьев поясницы коричневатым оттенком. НКХ (нижние кроющие хвоста) черные, с широкими белыми вершинами, остатками коричневых полос по бокам опахал и (не всегда) белыми наствольными полосами или пятнами в средней части пера.
Самец в первом взрослом наряде. Единственное отличие от взрослой птицы — окраска наружного опахала ПМП-10, несущего беловатый крапчатый рисунок (Ellison, 1968).
Взрослая самка.
Окраска верхней части тела пестрая, серо-коричневая, с неясным поперечным рисунком из широких серых (с черноватой струйчатостью) вершинных, черных предвершинных и расположенных ниже охристых полос. На всех перьях верхней части тела — один и тот же рисунок из охристых полос по черно-бурому фону. Серые вершинные полосы особенно широки на спине. Летом, при изнашивании этих полос, на поверхность выходят предвершинные черные полосы и верх птицы становится очень темным. На нижней части тела господствует правильный поперечно-полосатый рисунок из белых, черно-бурых и охристо-желтых полос. Белые вершинные полосы особенно широки на брюхе, нередко придавая ему почти белую окраску, и отсутствуют (или очень узки) в верхней части груди, которая имеет охристую окраску. Горло беловатое в бурых и охристых пестринах. Окраска перьев крыла, как и у самцов, но кроющие перья нередко имеют желтовато-коричневую окраску (встречающуюся у самцов очень редко) и белые наствольные полосы бывают на них гораздо чаще. РП черно-бурые, с неясным поперечным рисунком из тонких охристых полос и с широкими охристыми вершинными пятнами, нередко-испятнанными черной струйчатостью и не имеющими в отличие от самцов черной окантовки. На центральной паре, окрашенной под цвет поясницы и надхвостья, развит правильный поперечно-полосатый рисунок, но он также размыт желтовато-серой струйчатостью.
Ювенальный наряд (самец и самка).
Окраска почти такая же, как и у азиатской дикуши, только окраска груди более интенсивного охристого цвета, на внутренних ВМП лучше развит поперечный рисунок из черных полос, а перья поясницы охристые с правильными темно-бурыми поперечными полосами.
Окраска пуховых птенцов такая же, как и у азиатской дикуши, только черное окаймление ярко-коричневой «шапочки» на темени выражено гораздо лучше.
Размеры и масса.
Размеры вида указаны в таблице 1 статьи о дикушах. Масса взрослых птиц колеблется в пределах 460-580 г у самцов и 430-570 г у самок, испытывая сезонные колебания, типичные для тетеревиных птиц (Pendergast, Boag, 1973; рис. 3). Крыло несколько более узкое и острое, чем у азиатской и горной дикуш. Вершину крыла составляет либо ПМП-8, либо ПМП-7, иногда, оба вместе.
Распространение и ареал.
Ареал канадской дикуши охватывает всю таежную зону Северной Америки от тихоокеанского до атлантического побережья (рис. 4). По заросшим еловыми и смешанными лесами долинам рек этот вид довольно далеко проникает в лесотундру, и его северная граница довольно извилиста. Начинаясь немного севернее Полярного круга на побережье зал. Коцебу у устья р. Ноатак, она идет по лесистым долинам правых притоков Юкона, стекающих к югу с гор Эндикотт, на восток через лесистые долины Поркьюпайна и Олд-Кроу к низовьям р. Маккензи и р. Андерсон, через северо-восточные побережья Большого Медвежьего и Большого Невольничьего озер, долину р. Кокран и оз. Броше к устью р. Черчилл на западном берегу Гудзонова залива. На восточном берегу граница начинается от зал. Ричмонд и идет к северо-востоку, к южной окраине зал. Унгава (Форт-Чимо, Джордж-Ривер) и отсюда к восточному берегу Лабрадора, выходя к атлантическому побережью у о-вов Окак (Bent, 1932; Aldrich, 1963; Todd, 1963). Южная граница начинается на атлантическом побережье около 43° с. ш. и через южную часть штата Нью-Гемпшир, северную часть штатов Вермонт и Нью-Йорк идет к оз. Онтарио, обходя его с севера, пересекает озера Гурон и Мичиган примерно под 44°30 с. ш. и дальше через северную часть штатов Висконсин и Миннесота огибает с севера прерии Южной Канады, выходя к подножью Скалистых гор в юго-западной Альберте. Отсюда граница следует на северо-запад вдоль подножья Скалистых гор до истоков Смоки-ривер, где она углубляется в горы и идет к тихоокеанскому побережью, но к нему не выходит, идя вдоль восточных склонов гор Святого Ильи и северных склонов Аляскинского хребта, достигая берега Тихого океана в районе Бристольского залива. Положение границы ареала в Скалистых горах, где канадская дикуша инерградирует с горной дикушей выяснено крайне недостаточно. Вне очерченных таким образом границ сплошного ареала имеется еще один изолированный участок, занимающий прибрежные области у заливов Кука и Принс-Вильяма, южнее Аляскинского хребта. На востоке ареала канадская дикуша населяет о-ва Кейп-Бретон и Принца Эдуарда, на западе о-ва Кетчикан и Принца Уэлльского — все не далее 20 км от материкового берега. На более отдаленных от материка островах отсутствует, но недавно были попытки акклиматизации ее на о-вах Кадьяк (1957 г.) и Ньюфаундленд (1964 г.). Результаты этих попыток пока неизвестны (Long, 1981). Нет дикуши и на о-ве Ванкувер, хотя пролив, отделяющий его от континента, местами не превышает 6 км в ширину.
Места обитания.
Основные места обитания приурочены к темнохвойной тайге, где преобладающими породами являются ель, пихта и сосна и где почву, захламленную буреломом, одевает густой моховой ковер или заросли багульника, густые древостои сменяются открытыми пятнами болот с разреженной древесной растительностью. У северо-западной границы ареала дикуша выходит почти к самой границе леса, обитая по разреженным ельникам вдоль русел рек (Bent, 1932). В центральной части ареала, например в провинции Онтарио, дикуши обитают в сосновых лесах, где преобладает сосна Бангса (Pinus banksiana), с подростом из белой ели Picea glauca, перемежающихся с сырыми массивами черной ели P. mariana (Lumsden, 1961а). В штате Мичиган наиболее обычные места обитания — разреженные сосняки, разновозрастные ельники с примесью тополя, осины и березы или же болота с разреженным древостоем из черной ели и лиственницы, чередующиеся с приподнятыми гривами, заросшими сосняками (Amman, 1963). На Аляске основными местами обитания являются еловые леса с преобладанием черной ели в низинах и смешанных древостоев из белой ели, березы, осины и тополя на возвышенных местах (Ellison, 1975).
Численность.
Гнездовая численность на юге заметно выше, чем на севере, и составляет, например, в штате Мичиган 156-196 птиц на 1000 га (Robinson, 1969), а на Аляске только 90-123 птицы на 1000 га (Ellison, 1975). Осенняя численность не подсчитывалась, но, судя по приведенным данным, она может составлять до 500 птиц на 1000 га.
Популяционный состав.
На Аляске при всех осенних учетах соотношение самцов и самок было одинаковым и только раз равновесие было нарушено — на 100 самок приходился 61 самец (Ellison, 1974). Соотношение самок, самцов и молодых птиц по результатам осенней охоты в провинции Онтарио, Канада, менялось по годам в значительных пределах. За одним исключением среди взрослых птиц преобладали самцы (от 104 до 168 самцов на 100 самок), тогда как среди молодых птиц в течение 4 лет из 6 преобладали самки (85-92 самца на 100 самок), в одном случае соотношение было почти равным (100:103) и в одном преобладали самцы (122 на 100 самок). Число молодых птиц на взрослую самку колебалось от 2.5 до 7.3 (Lumsden, Weeden, 1963).
Территориальность.
Канадская дикуша отличается высокой оседлостью. При мечении птиц в штате Мичиган, США, у трех самок среднее удаление от места кольцевания составило за год 2.6 км, а у 8 самцов — 1.4 км, причем максимальная удаленность не превышала 5.6 км (Robinson, Maxwell, 1968). На Аляске перемещения молодых птиц в первую осень составляют в среднем 3.2 км (минимальное 0.2 и максимальное — 10.5 км). Средняя величина удаления взрослых птиц (27 особей) от места поимки составила за год менее чем 1 км. Птицы не уходили далее чем 1.6 км, и только раз одна из них откочевала на 8 км. Осенние перемещения вообще наиболее значительны в течение года, что связано в основном с поисками гальки. В целом же индивидуальная территория взрослой птицы в зависимости от места обитания может занимать от 18.8 до 177 га. Зимние перемещения исследованы мало. В одном случае самка провела 2 месяца в пределах 3 га, в другом — самец в течение ноября-декабря перемещался по площади около 102 га (Ellison, 1973). Пространственная структура популяции в период размножения состоит из токовых участков самцов, располагающихся обычно группами. Строгий территориализм свойствен только взрослым самцам и только в период размножения. Наблюдения с помощью радиопередатчика позволили установить, что взрослый самец занимает сравнительно большой участок — от 1 до 3.5 га. Только в очень редких случаях молодые самцы также могут обладать территорией и размеры токовых участков при этом могут быть очень большими, до 8 га. Обычно же молодые самцы свободно перемещаются между участками взрослых птиц и ведут кочевую жизнь. В одном случае такой молодой самец переместился за сутки на 1.6 км. Привязанность к токовому участку у самцов резко ослабевает в конце мая, когда самки приступают к насиживанию и индивидуальная территория самца намного увеличивается. Типичным можно считать поведение самца, территория которого между 21 апреля и 21 мая была равна примерно 1 га, а к 22 июля самец бродил по территории в 6 раз большей (Ellison, 1971).
Географическая изменчивость.
Географическая изменчивость выражена незначительно благодаря сравнительно однообразным условиям обитания на протяжении всей таежной полосы Северной Америки. Только на юго-западе и юго-востоке ареала, где эти условия заметно отличаются большей мягкостью и влажностью, а также более богатой растительностью, обитают две формы, вполне отличимые по окраске от занимающего основную часть ареала номинативного подвида, — F. c. atratus и F. c. canace.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные.", 1985 г.
 Азиатская дикуша. Falcipennis falcipennis.
Азиатская дикуша. Falcipennis falcipennis. Миноги.
Миноги.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Дикуши. Род Falcipennis.
Дикуши. Род Falcipennis..") Обыкновенный рябчик (Bonasa bonasia).
Обыкновенный рябчик (Bonasa bonasia). sewerzowi.") Рябчик Северцова. Bonasa (Tetrastes) sewerzowi.
Рябчик Северцова. Bonasa (Tetrastes) sewerzowi. Воротничковый рябчик. Bonasa umbellus.
Воротничковый рябчик. Bonasa umbellus. Рябчики.
Рябчики.
