Обыкновенный рябчик (Bonasa bonasia).
Лесная птица средних размеров, со сравнительно коротким закругленным крылом, хвостом умеренной длины и пестро окрашенным оперением с преобладанием серого и темно-бурого цветов. Половой диморфизм в окраске и размерах незначителен.

Рябчики.
Окраска самца.
Окраска самки.
Ювенальный наряд.
Пуховый наряд.
Размеры и масса рябчиков.
Строение тела.
Линька.
Ареал рябчиков и их распространение.
Места обитания.
Смертность и численность.
Численное соотношение полов рябчиков.
Территориальность.
Образ жизни.
Повадки и образ жизни в теплое время года (весна-лето).
Образ жизни зимой.
Питание.
Токование и размножение.
Гнездование.
Охрана и хозяйственное значение.
Географическая изменчивость.

Самец рябчика.
Окрас оперения взрослого самца.

Птенец рябчика.
Верх головы, шеи и передняя часть спины серые, с рыжеватым оттенком и поперечным рисунком из частых темно-бурых полос, толщина которых варьирует и они могут утоньшаться настолько, что образуют струйчатый рисунок. Эти поперечные полосы формируются предвершинными полосками каждого пера, основной же серый фон образуют вершинные полосы. Все перья на этих частях тела окрашены однотипно, с чередованием темных и светлых полос. Верхняя полоса серовато-белесая, предвершинная темно-бурая, следующая — опять серая, затем опять бурая, опять серая и наконец бурое основание. Число полос варьирует от 5 до 7. Остальная часть спины, поясница и надхвостье серые или светло-серые с неясным поперечным рисунком из светлых полосок и редких, но четких продольных штрихов бурого цвета. Серый фон образуют те же самые вершинные полосы перьев, которые здесь более широкие и на самом конце пера имеют беловатую каемку — она и дает неясный поперечный рисунок. Предвершинные же бурые полосы теряют монолитность, дробятся и часто образуют тонкий струйчатый рисунок, иногда выходящий на поверхность оперения. Основания перьев бурого цвета, и по стержню пера этот бурый цвет выходит к вершине заостряющейся полоской — они и образуют продольные штрихи.

Птенец обыкновенного рябчика из Сибири.
Специфичный рисунок самца особенно хорошо выражен на горле и груди. Горловое черное пятно окружает широкая белая полоса, идущая ото лба под глаз и далее вниз. По центру лба эта полоса прерывается, четко оторачивая сзади черную окраску ноздрей. Контрастность белой полосы подчеркивает черное окаймление ее с наружной стороны, особенно по бокам головы и на лбу. Бока головы имеют более рыжую окраску, позади глаза располагаются два белых пятна. Еще одна белая полоса расположена по бокам шеи у ее основания и идет назад и вниз, к зашейку. Ее формируют белые вершины перьев, ограничивающих шейную аптерию с дорсальной стороны. Во время токования самец, меняя положение перьев, может менять конфигурацию и размеры этих полос и пятен.
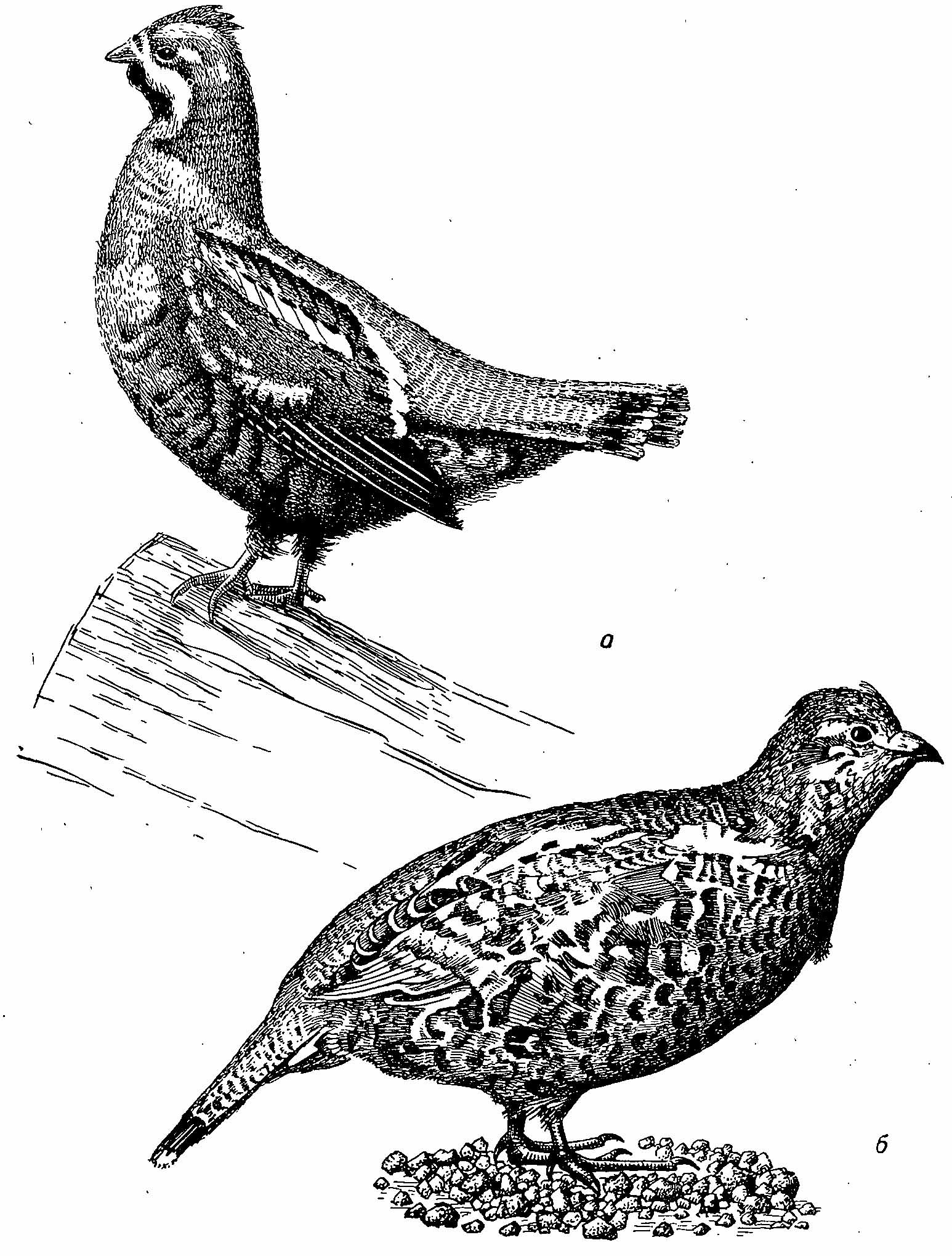
Рис. 1. Обыкновенный рябчик Bonasa bonasia. а — самец; б — самка.
Нижняя часть тела окрашена более пестро, рисунок здесь крупнее и формируется в основном белыми вершинами перьев. Наиболее темно окрашена область зоба, где серовато-коричневый фон, нередко с рыжеватым оттенком, испещрен правильным рисунком из подковообразных белых полосок. Далее назад эти полоски становятся шире, сливаются между собой и вместе с широкими белыми вершинами образуют преимущественно белую окраску брюха, по которой равномерно разбросаны темные пятна. По бокам груди, ниже основания крыльев, расположены партии перьев с интенсивно-рыжей окраской и белыми вершинами, предваренными черной полосой. В обычной позе эти партии перьев скрыты сложенным крылом. На перьях зоба по вершине каждого пера идет тонкая черноватая полоска, плохо заметная на темном фоне следующего пера. За ней расположена широкая белая полоса характерной скобковидной формы (рис. 2). За ней перо окрашено в черно-бурый цвет, постепенно светлеющий к основанию пера. На задних партиях перьев и по бокам груди есть еще зачаток второй белой полоски в середине пера, имеющей вид наствольного пятна, нередко смещенного на одно из опахал. По направлению к брюху вершинная черная каемка исчезает, и белая полоса становится вершинной, одновременно увеличиваясь в размерах. Остальная же часть пера приобретает поперечно-полосатую окраску, причем вторая белая полоса по краям опахала обычно соединяется с первой, а предвершинная темная полоса принимает дугообразную форму и образует на брюхе характерный чешуйчатый узор. Сильно рассученные перья на задней части брюха двуцветны — с буровато-серым основанием и белой вершиной. Длинные перья боков тела также поперечно-полосатые, часто они окрашены асимметрично, и наружное опахало теряет рисунок, становясь одноцветным кирпично-бурым. Окраска крыла пестрая, с белыми и бурыми пятнами по серому или серовато-охристому фону. Здесь выделяется прежде всего широкая белая полоса, идущая вдоль спины по плечевым перьям и как бы продолжающая шейную полосу. Вторая светлая полоса идет по заднему краю крыла, по вершинам второстепенных маховых перьев. Мелкие белые пятна по поверхности крыла — это вершины многих больших, средних и малых кроющих перьев. Пятна эти имеют характерную треугольную форму, вершиной к основанию пера. Окраска кроющих перьев крыла характерна асимметричным рисунком, причем на одних перьях поперечные полосы исчезают на наружных опахалах (проксимальные средние кроющие, малые кроющие), у других — на внутренних (большие кроющие, дистальные средние кроющие). Наиболее сложный узор несут вершинные части проксимального ряда плечевых и внутренних ВМП (второстепенные маховые перья). У последних в верхней части внутреннего опахала темные полосы преобразованы в черно-бурые пятна сложной формы, вершинная же белесая полоса получает большое распространение на внешнем опахале (рис. 3). В результате на сложенном крыле по его заднему краю образуется беловатая полоса, которую предваряет ряд бурых продольных штрихов — выступающих из-под налегающих перьев пятен внутренних опахал. ПМП (первостепенные маховые перья) и ВМП буровато-серые, с желтоватыми поперечными полосками на внешнем опахале, ВМП с белой вершинной окантовкой. У наружных ПМП внешние опахала становятся совсем светлыми, полосатый рисунок на них редуцируется почти полностью.
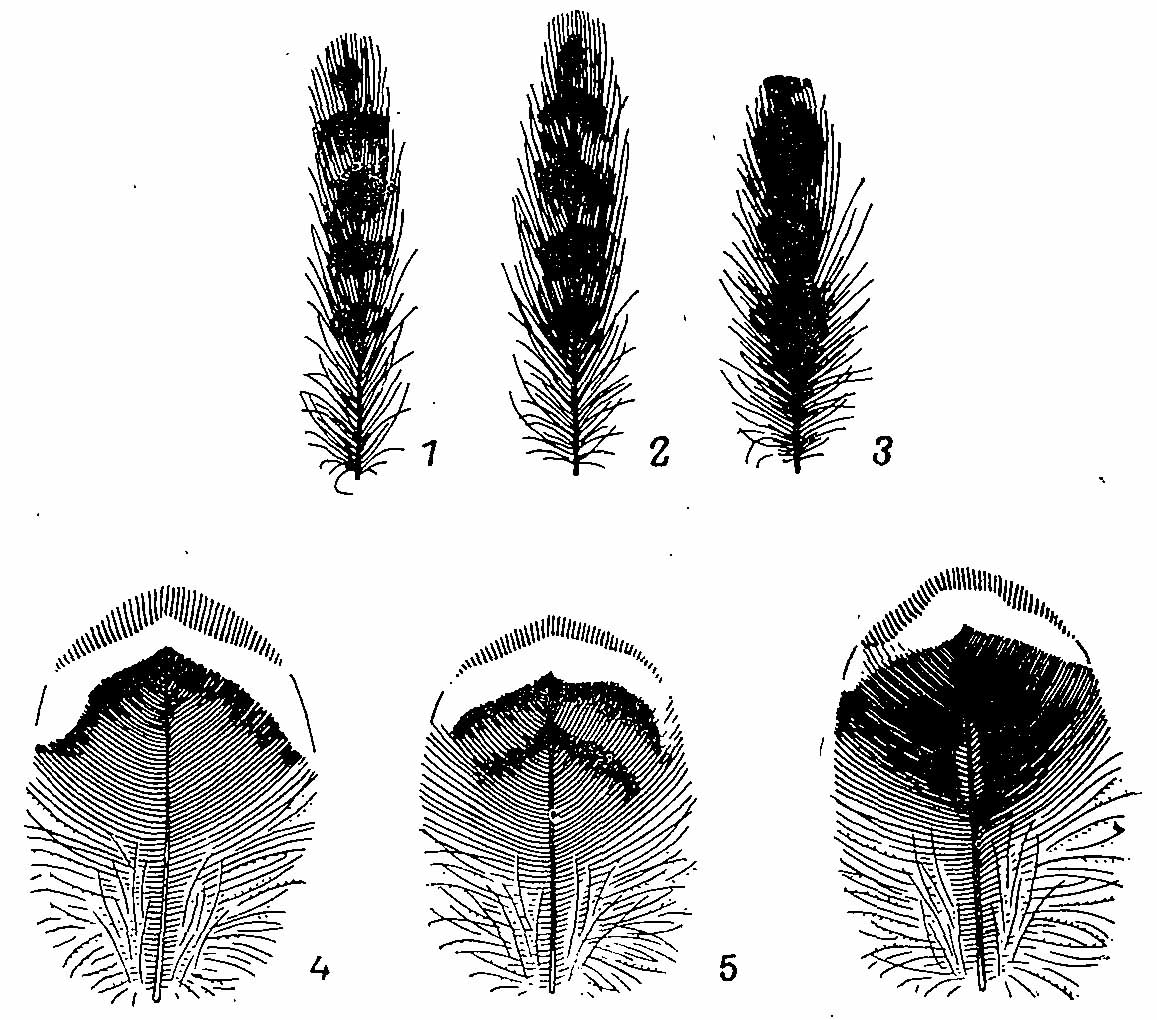
Рис. 2. Перья с хохолка (1—3) и с центра груди (4—6) Bonasa bonasia. l, 4 — взрослый самец; 2, 5 — молодой самец; 3, 6 — взрослая самка.
Окраска РП (рулевых перьев) поперечно-полосатая, с широкой предвершинной черной каймой. Узкая вершинная полоса серовато-желтая, нередко с остатком струйчатого рисунка. Остальная часть РП — чередование более узких, разбавленных струйчатостью светло-серых и буровато-охристых полос. Центральная пара (иногда даже две центральные пары) резко отличается по окраске от остальных — без черной предвершинной полосы, поперечно-полосатый рисунок вообще выражен очень нечетко, будучи затушеван сплошным струйчатым узором по серому фону. По общему тону окраски эти перья не отличаются от спины и при сложенном хвосте прикрывают более контрастную, демаскирующую окраску остальных РП. НКХ (нижние кроющие перья хвоста) с поперечно-полосатым рисунком и широкими белыми вершинами.
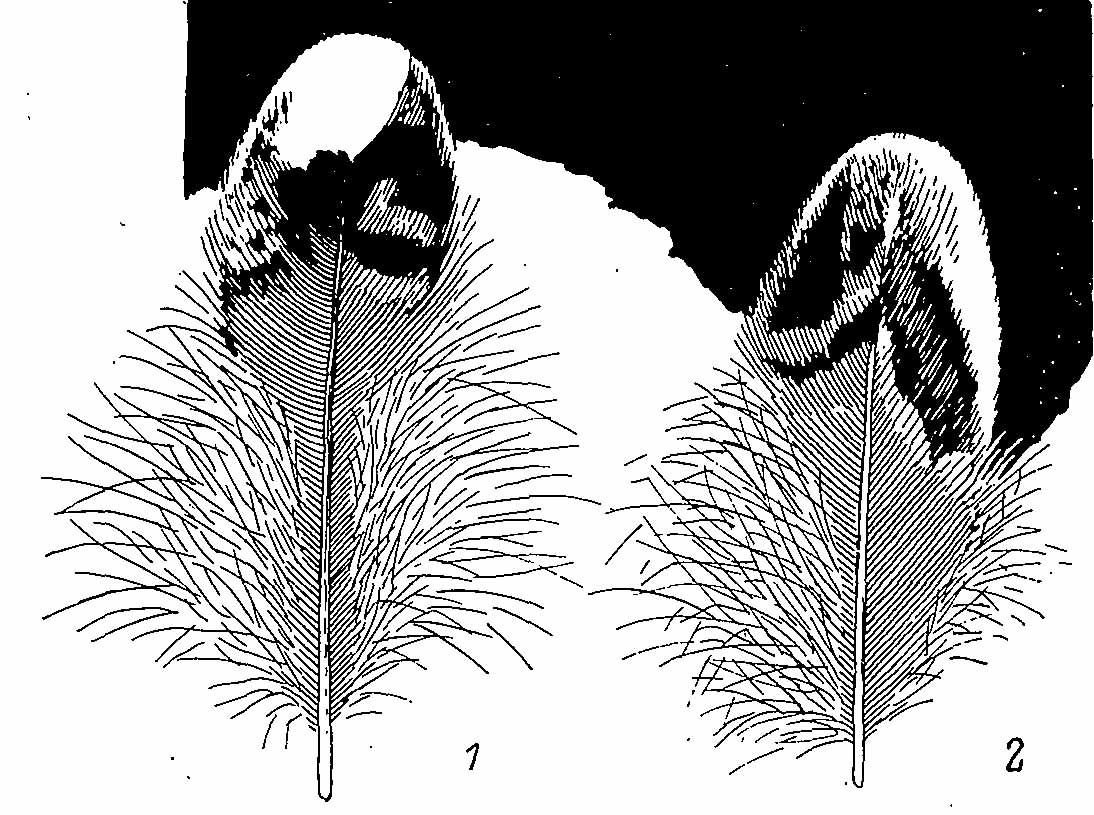
Рис. 3. Внутреннее маховое перо (1) и плечевое (2) перо самца Bonasa bonasia.
Самец в первом взрослом наряде.
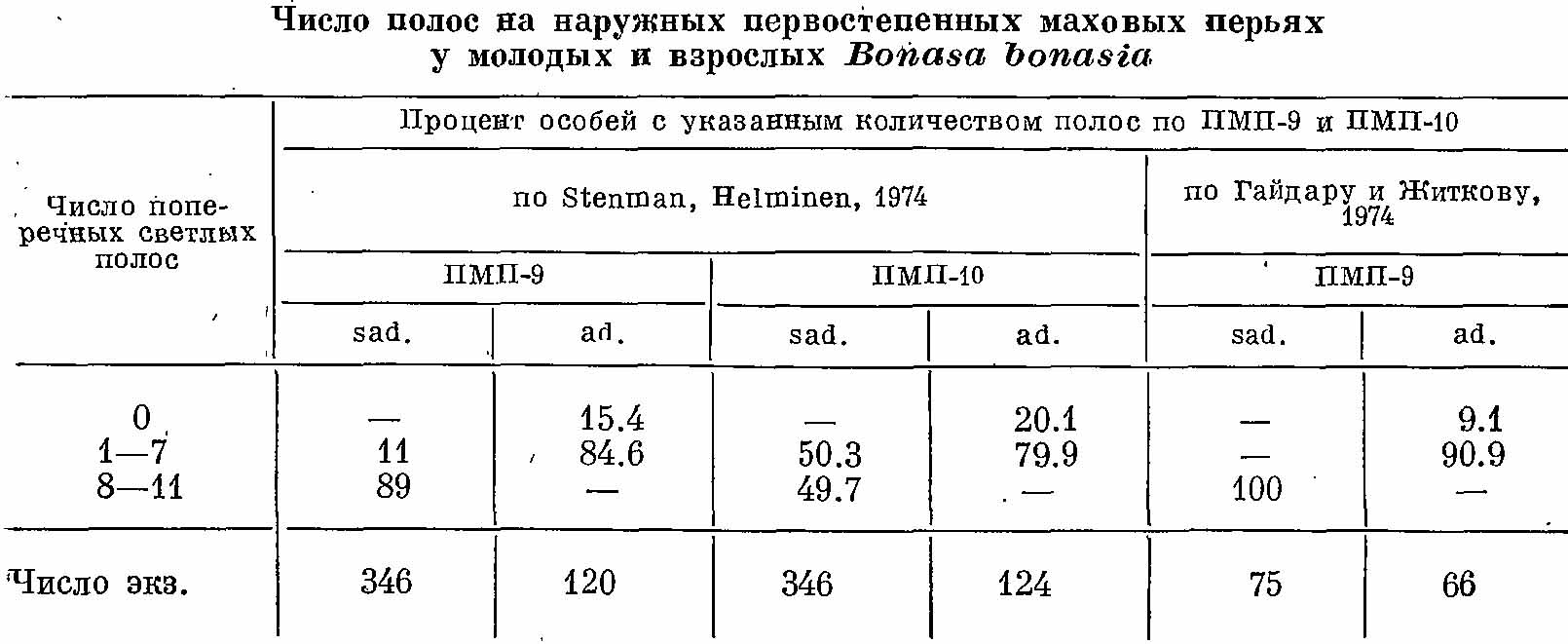
Таблица 1. Число полос на наружных первостепенных маховых перьях у молодых и взрослых Bonasa bonasia.
Отличается от окончательного несколько большей диффузностью окраски и более развитой поперечной полосатостью рисунка — прежде всего на наружных опахалах ПМП-9 и ПМП-10, а также на перьях пластрона. Наружные опахала упомянутых маховых перьев несут большое количество поперечных полос (табл. 1). Кроме того, ПМП-1 имеет нечетко ограниченную светлую вершину с мелким крапчатым рисунком — в окончательном наряде здесь имеется только светлая каемка. На передней части грудного пластрона перья несут поперечно-полосатый рисунок (ниже белой предвершинной полосы) или ромбовидные пристержневые пятна желтовато-охристого цвета.
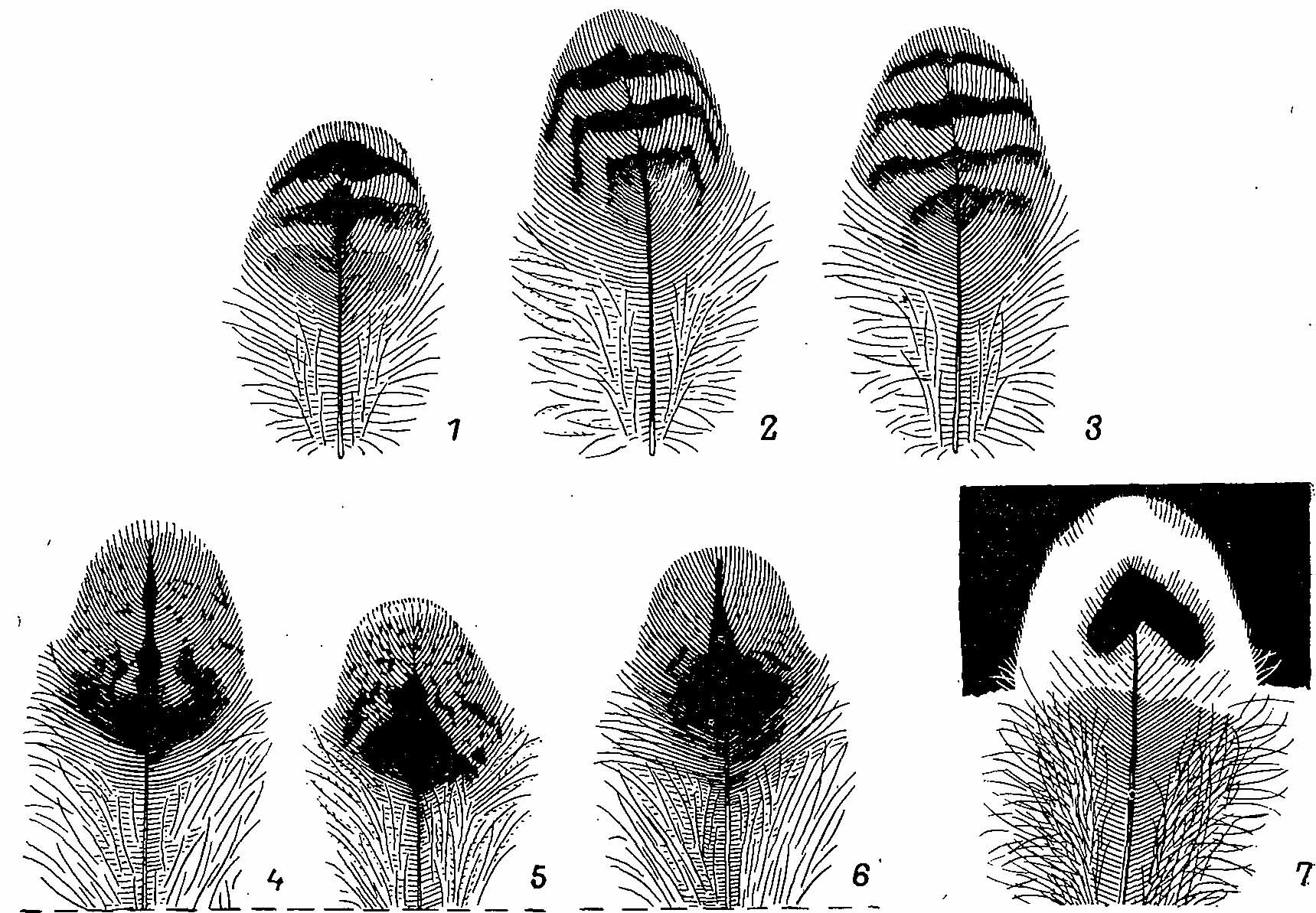
Рис. 4. Перья со спины (1—3), с поясницы (4—6) и перо с нижней части груди (7) Bonasa bonasia. 1,4 — взрослый самец; 2,5 — молодой самец; 3,6,7 — взрослая самка.
Взрослый самец в летнем наряде (июль).
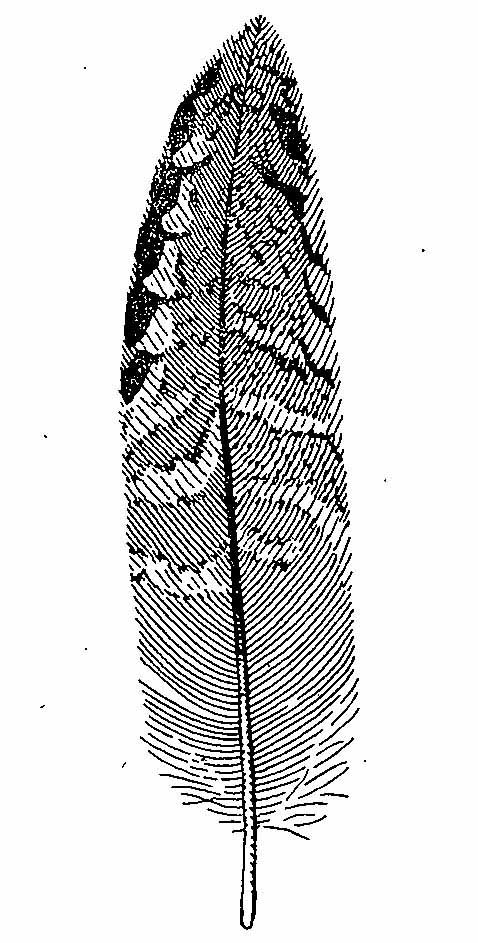
Рис. 4.1. Внутреннее маховое перо ювенального наряда Bonasa bonasia.
На боках головы и вокруг горлового пятна вырастает ряд мелких перьев с примитивной пестрой окраской, благодаря которым белое окаймление горлового пятна почти теряется в бурых пестринах. У южных популяций такие перья появляются и в нижней части горлового пятна. Кроме того, по бокам шеи и головы есть отдельные перья с мелкими каплевидными пятнами на концах, как у ювенальных.
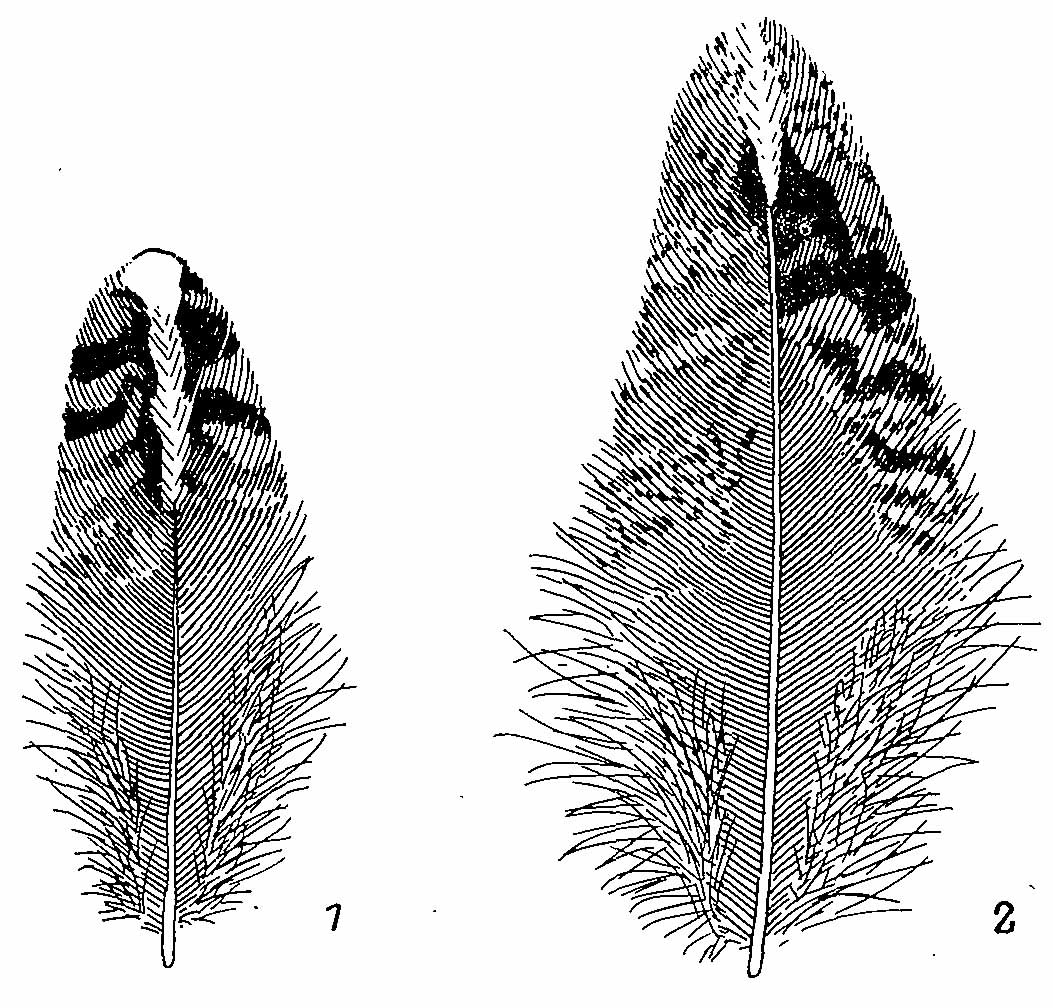
Рис. 5. Перья ювенального наряда Bonasa bonasia. 1 — со спины; 2 — плечевое.
Окраска перьев взрослой самки.
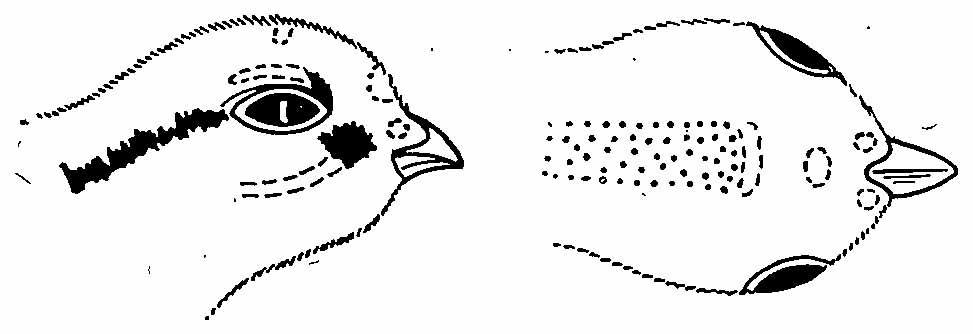
Рис. 6. Рисунок на голове пухового птенца Bonasa bonasia. Прерывистая линия — места, где черный цвет развит не у всех особей; точки — коричневый цвет.
Окраска оперения еще более пестрая, чем у самцов. Горловое пятно только намечается отдельными бурыми пятнами но сероватому фону. Эти пятна могут располагаться очень густо и в некоторых случаях даже способны образовать сплошное черновато-бурое горловое пятно, как и у самцов, однако яркой черной окраски, свойственной последним, оно никогда не имеет. В окраске перьев темени, зашейка, передней части спины в отличие от правильного поперечно-полосатого рисунка самцов преобладают различных размеров бурые пятна, а сам общий фон с заметным рыжим оттенком. Эти пятна формируются предвершинными полосами перьев, которые более широкие, чем у самцов, и нередко сливаются со второй бурой полосой (рис. 4). Задняя часть спины, поясницы и надхвостье обычно не имеют такого чистого серого фона, как у самцов, главным образом благодаря бурому струйчатому рисунку на вершинной части перьев. Окраска перьев груди и хохолка также более примитивна (рис. 2).
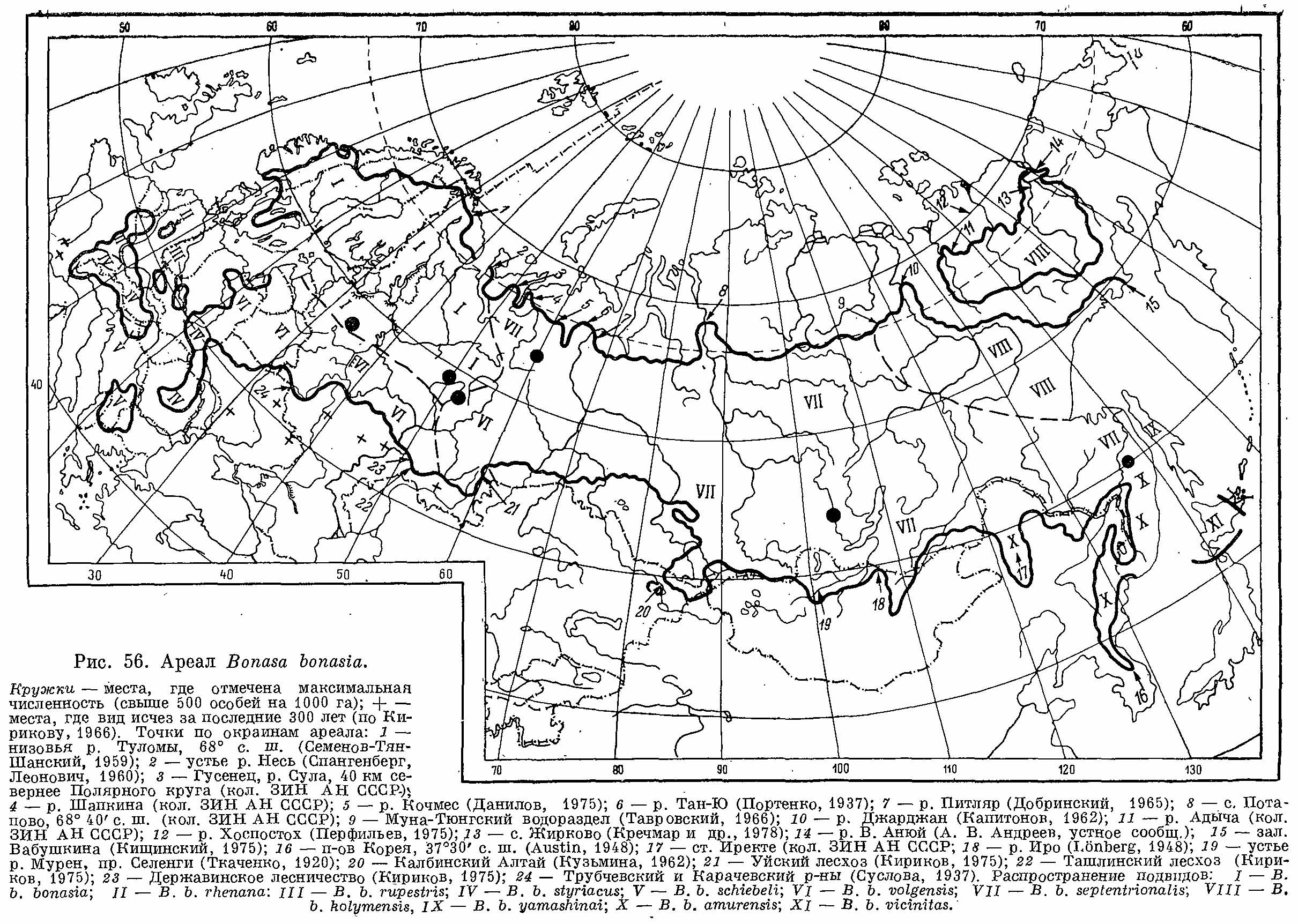
Рис. 7. Ареал рябчиков обыкновенных (Bonasa Bonasia).
Самка в первом взрослом наряде.

Таблица 2. Численность Bonasa bonasia на 100 га в обычных местах обитания.
Основные отличия от окончательного взрослого наряда сводятся к тем же особенностям окраски внешних опахал ПМП-9 и ПМП-10, что и у молодых самцов. У некоторых особей на нижней части шеи и на верхней части груди много перьев с белыми каплевидными пятнами у вершины. Такое пятно обычно бывает расположено в центре предвершинной охристой полосы.
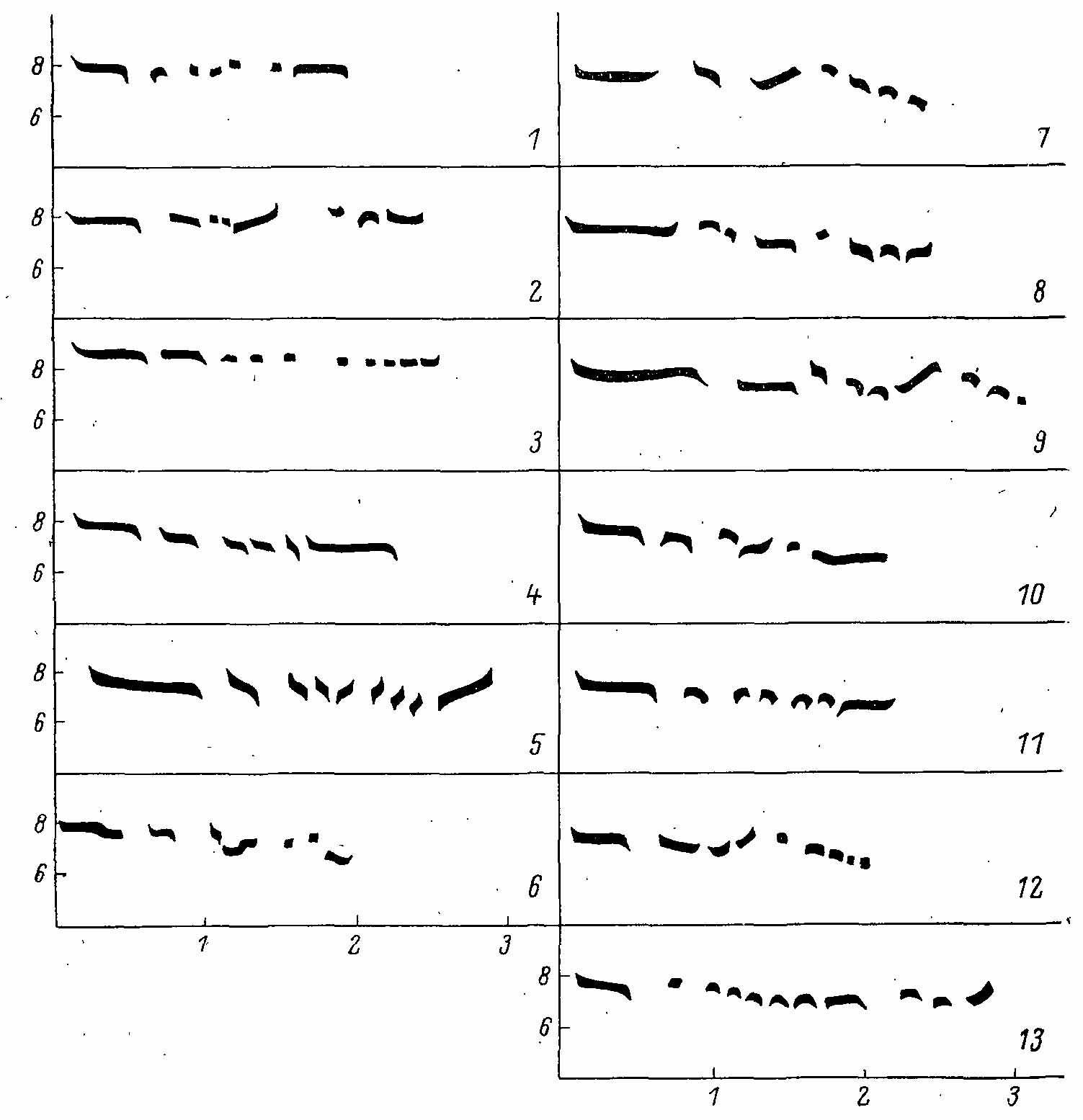
Рис. 8. Графическое изображение уровня и длительности свистовых нот песни Bonasa bоnasia из различных мест Европы.
Ювенальный наряд.

Рис. 9. Токовые позы Bonasa bonasia (по фотографиям).
Верхняя часть тела от темени до лопаток пестро окрашена в красновато-коричневый и темно-бурый цвета. На этом фоне разбросаны узкие белые штрихи — наствольные полоски, выходящие к вершине пера, но большая часть перьев таких полосок не имеет. Перья в этой области окрашены по тому же типу, что и у взрослых птиц, но только вершинная полоса не серая, а охристая. Спина, поясница и надхвостье окрашены более однотонно — неясные поперечные буроватые полоски по серому фону. Каждое перо имеет несколько таких полосок, которые нередко дробятся и дают струйчатый рисунок. На темени имеются удлиненные перья, образующие подобие хохолка. На боках головы намечается заглазничная белая полоса. Низ тела окрашен более однородно. На общем светлом фоне выделяется только пестрая окраска груди, где перья несут довольно разнообразный рисунок, в котором едва намечается поперечная полосатость. На многих перьях вершинной белой полосы нет, а есть только белое каплевидное пятно. РП серовато-бурые, центральная пара с рыжеватым оттенком. Поперечно-полосатый рисунок хорошо заметен в основной части пера и едва намечен в верхней его половине. Для перьев крыла свойственны белые каплевидные пятна на вершинах и резкая асимметрия рисунка, образуемого черно-бурыми полосами, которые, сливаясь, дают довольно сложный узор. ПМП несут четкий поперечно-полосатый рисунок на наружных опахалах. Внутреннее опахало более однообразной буроватой окраски. На наружных ВМП — нечеткие поперечные полосы, на внутренних же вдоль внешнего края идет своеобразный зубчатый рисунок черно-бурого цвета (рис. 4.1, 5) — очень характерный для ювенального наряда данного вида.
Пуховой наряд.
Верх головы и шеи, спина, поясница и надхвостье коричневые, нередко с охристым оттенком. Лоб желтовато-рыжий. Бока головы и нижняя часть тела серовато-желтые, зоб иногда с охристым оттенком. Рисунок на голове образован надглазничной (иногда бывает очень короткой), заглазничной и нижнечелюстной (нередко отсутствует) полосами, а также пятном перед глазом (рис. 6). У отдельных экземпляров бывают еще пятна на ноздрях и в основании конька надклювья.
Как видно из этих описаний, развитие окраски самца рябчика в онтогенезе сводится к более четкой локализации намеченных в ювенальном наряде отличий в окраске зоба, горла, брюха, передней и задней половин верхней части тела.
Значительно более резким становится контрастный рисунок головы и шеи — горло делается сплошь черным, обрамленным широкой белой полосой. Белые полосы появляются по бокам шеи, более резко выделяется заглазничное белое пятно. На перьях спицы исчезают белые клиновидные полосы, сохраняясь только на перьях крыла. На перьях зоба первичная полосатость сменяется специфичной окраской. Более контрастной становится и окраска хвоста, прежде всего за счет появления здесь широкой черной предвершинной полосы.
У самок изменений в процессе онтогенеза меньше. Горло за редкими исключениями остается пятнистым, хотя и в меньшей степени, чем у молодых птиц. Белая окантовка горла и белые полосы на шее также менее четкие, чем у самцов. На перьях груди сохраняется поперечно-полосатый рисунок.
Размеры и масса.
Значительные колебания в размерах обусловлены возрастной, половой и географической изменчивостью. Ниже даются следующие размеры коллекционных экземпляров ЗИН АН СССР (номинативный подвид) в миллиметрах.
Самцы (28 штук): крыло 167.6 (средняя величина) (162—180), хвост 113.5 (104—122), длина клюва (от переднего края ноздри) 10.4 (9.3—12.1), плюсна 33.5 (28—36).
Самки (18): крыло 166.6 (162—171), хвост 107.2 (100—115), длина клюва 10.9 (10—12.3), плюсна 33.3 (32—35).
Масса заметно меняется по сезонам, будучи максимальной поздней осенью и в начале зимы. К весне она постепенно снижается и у самцов становится минимальной в конце весны, хотя по некоторым данным (Волков, 1975), она поднимается вновь в марте на 18 % с момента появления проталин. Аналогично кратковременное увеличение массы весной отмечено и для B. umbellus (Bump et al., 1947). У самок масса после зимнего снижения вновь заметно возрастает к началу насиживания и затем падает до минимальных величин во время появления птенцов. Масса самцов колеблется по всему ареалу и по разным сезонам от 317 до 580 г, самок — от 305.5 до 560 г.
Строение тела.
Крыло закругленное, тупое. Его вершину образуют 7-е или 7-е и 6-е первостепенные маховые, формула крыла — 7>6>8>5>9>4. Хвост слегка закруглен и крайние перья на 7—9 % короче центральных. Плюсна оперена на 2/3 длины, у северо-восточных популяций — на ,3/4 длины и даже (у отдельных экземпляров) до самого основания. По бокам пальцев на зиму отрастают роговые зубчики длиной до 3.5 мм. Они также максимально развиты у северных и северо-восточных популяций. Клюв типично тетеревиного типа, короткий, толстый, слегка загнутый вниз, с режущими краями рамфотеки. «Бровь» у самца очень мала и вне периода токования совсем незаметна. Цвет радужины бурый, ног — серый, клюва — черноватый.
Линька.
Последовательность смены нарядов — обычная для тетеревиных птиц: пуховой—птенцовый—первый дефинитивный—первый летний—второй дефинитивный (окончательный)—второй летний и т. д. Первый наряд целиком пуховой, только на крыльях в первые же сутки появляются кисточки 7 ПМП и 10 ВМП. Первая линька постнатальная, полная и приводит к замене пухового наряда перьевым ювенальным, хотя пух предшествует не всем перьям. Без пухового предшественника растут те перья, которые продуцируют поздно созревающие фолликулы, расположенные на спине по окраинам птерилий. В процессе этой линьки сначала развиваются маховые перья (за исключением трех наружных), далее появляются перья на спине и боках, затем на брюшке и потом уже на голове и шее. В первый же этап линьки на голове вырастают перья хохолка, а на самом последнем — ПМП-8. Постювенальная линька начинается заменой маховых перьев на 16-й или 17-й день жизни. Смена перьев — типичная для тетеревиных. Почти одновременно с началом смены ювенальных маховых перьев дефинитивными начинают расти ПМП-9, а затем и ПМП-10 первой генерации. Таким образом, крыло молодой птицы несет перья двух генераций, где к первой относятся два наружных ПМП, а также внутреннее перо крылышка. Форма и окраска этих перьев, как уже говорилось, позволяют достаточно надежно отличать молодую птицу от старой. Несколько позже маховых линяют рулевые и контурные перья. Последние начинают сменяться сначала на спине, затем и на брюхе. У самцов черный цвет на горле появляется сначала полоской на самой середине подбородка, затем двумя полосами по краям горла. Вслед за этой «трехполосной» стадией сменяются и остальные перья горлового пятна. Очередная полная линька начинается следующим летом в конце мая—начале июня с замены ПМП-1. На нее в июне—июле накладывается частичная летняя линька, которая охватывает в основном переднюю часть тела — голову, шею, зоб, переднюю часть спины и груди. Линяют также отдельные перья на боках и спине до надхвостья включительно. У северных популяций летняя линька выражена слабо, летние перья появляются только на голове (в основном на подбородке) и вентральной стороне шеи — все это происходит с начала июля.
Самцы начинают линять в последние дни токования, хотя основная летняя линька происходит после окончания тока. С апреля у птиц начинают отпадать роговые бахромки на пальцах, и они полностью исчезают к концу мая. У самок летняя линька происходит в более сжатые сроки и оканчивается к периоду насиживания. При летней линьке, как уже говорилось, часть выпавших перьев новыми не заменяется, т. е. происходит разреживание оперения. Летний наряд носится недолго, около месяца или немногим более. У самцов летняя линька под конец смешивается с осенней и переходит в нее, у самок же она достаточно четко отделена от осенней смены контурного оперения периодом насиживания и вождения птенцов, когда линька практически прекращается.
Сравнительно поздно, в конце августа, у самцов линяет горловое пятно. Предварительно часть черных перьев в нижней части пятна заменяется на летние перья, пестро окрашенные (бурые с белой вершиной), затем эта область линяет по той же схеме (с трехполосной стадией), что и у молодых птиц, но выпадающие перья заменяются новыми не сразу и во время линьки на горле появляются участки голой кожи.
Окончательный взрослый наряд надевается во время второй осенней линьки. И у взрослых, и у молодых она начинается с выпадания ПМП-1 и протекает в 2 фазы. Первая — основная, во вторую происходит только линька некоторых ВКК (верхние кроющие перья крыла), нижней части груди, брюха, а также дорастают маховые перья. Заканчивается эта линька к ноябрю.
Распространение.
Ареал вида охватывает почти всю лесную зону Евразии (рис. 7). Северная граница ареала определяется пределом распространения сплошных лесов. В Скандинавии распространен к северу до Скандинавских гор и только в одном месте выходит к морскому побережью. На Кольском п-ове дальше всего на север идет по долине р. Туломы (68° с. ш.), но далее на восток граница опускается к югу, обходя Хибины. Крайние точки на карте ареала дают представление о том, насколько далеко этот вид проникает на север. Дальше всего, за Полярный круг он идет по долине р. Шапкина в бассейне Печоры (кол. ЗИН АН СССР); по долине Енисея — до 68°40' с. ш. (с. Потапово, кол. ЗИН АН СССР), по долине Лены — до лесов долины р. Джарджан (69° с. ш.) и в междуречье Яны и Индигирки — до Адычи (68° с. ш.) и Хоспостоха (69°30' с. ш., Перфильев, 1975). Северо-восточная часть ареала представлена рядом изолированных участков по долинам крупных рек к северо-востоку от Верхоянского хребта. Восточная граница ареала идет по материковому побережью, захватывая также острова Сахалин и Хоккайдо. На востоке наиболее далеко к югу рябчик проникает в горах северной части п-ова Корея (37°30' с. ш.), откуда граница идет на запад, захватывая горный массив Джангуанцайлин в северо-восточном Китае, огибает с севера депрессию р. Сунгари, захватывает горы Малого Хингана и вновь опускается к югу по горам Большого Хингана до 47° с. ш. Отсюда граница вновь заходит на территорию СССР в верховьях р. Борзя и, огибая с севера борзинские степи, идет в Монголию, где вид этот населяет горные леса Хэнтэя и Хангая. От последнего граница идет по хр. Танну-Ола, охватывает леса Алтая (изолированный участок — на левом берегу р. Иртыш, в Калбинском Алтае) и поднимается на север вдоль западного склона Салаирского кряжа к лесным массивам Васюганья. Далее к западу южная граница ареала очень извилиста и не везде ее современное положение точно известно. Можно сказать только, что повсюду рябчик распространен до южных границ лесной зоны, проникая местами и в лесостепь, особенно по уремным зарослям ряда рек. В западной части СССР южная граница в последнее время постепенно смещается на север под воздействием хозяйственной деятельности человека, истребляющей или изменяющей леса. Особенно это характерно для западноевропейской части ареала. Сплошной ареал не распространяется западнее лесных районов восточной Польши, а дальше представлен изолированными участками, в основном приуроченными к лучше сохранившимся горным лесам Карпат, Альп, Динарид, Родоп, Старой Планины, Юры, Арденн, Вогез, Шварцвальда. Примечательно отсутствие рябчика в изолированных лесах Камчатки, Тянь-Шаня и Кавказа. Собранные мною сведения не подтверждают якобы недавнего обитания рябчика на Пиренеях. В коллекционных собраниях Франции и Великобритании экземпляры с Пиренеев отсутствуют. Возможно, начало этому заблуждению положила работа Бриссона (Brisson,, 1760), где «пиренейским рябчиком» назван белобрюхий рябок. Достоверны только ископаемые остатки этого вида, датируемые Мадленом (Couturier, 1964), а сообщения о недавних встречах его в лесах Пиренеев (Hartert, 1922) не подтверждены никаким документальным материалом.
Места обитания.
Будучи сугубо лесной птицей, рябчики могут населять самые различные типы леса, но особое предпочтение оказывают густым смешанным лесам по долинам речек, ручьев, на склонах оврагов и гор. Наиболее предпочитаются те типы леса, где в древостое преобладают береза, ель, ольха, а почва хорошо увлажнена и завалена валежником. Особенно тесно связаны рябчики с прирусловой растительностью в горах, а по северной окраине тайги и в лесотундровых редколесьях он живет только в пойменных зарослях. Явно избегаются чистые разреженные леса из сосны или лиственницы, а также сфагновые сосняки на верховых болотах. Только в южных частях ареала рябчик живет иногда в сосновых борах, там, где есть густые заросли папоротников. Столь же характерна явная приверженность к опушкам, краям просек или лесных дорожек, а также к местам контакта различных типов леса, особенно ельников и сосняков. Тесная связь рябчика с елью (Формозов, 1934; Донауров, 1947) наблюдается далеко не везде. На Карпатах они в ряде мест живут в буковых лесах (Портенко, 1950), на Южном Урале — в черемуховой урёме (Кириков, 1952), в Уссурийском крае — в пойменных зарослях из черемухи и дикого винограда (Белопольский, 1955). Северо-восточная часть ареала вида лежит вообще вне пределов распространения елей и пихт, и на Омолоне, например, рябчик живет в пойменной урёме из ивняков, ольхи, шиповника и лиственницы: (Кречмар и др., 1978).
Численность и смертность.
В оптимальных местах обитания численность вида может достигать осенью 120 птиц на 1 км2 (табл. 2), хотя повсюду она подвержена сильным колебаниям. Последние приобретают временами даже циклический характер, хотя эти циклы на различных участках ареала могут и не совпадать. Решающее влияние на численность оказывает климат и, в частности, условия погоды в момент массового вылупления птенцов и в течение первых трех недель их жизни. Так, например, коэффициент корреляции между осенней численностью рябчика и среднесуточными температурами мая—июня на Среднем Урале оказался наиболее высоким для второй и третьей декад июня, когда происходит массовое вылупление птенцов (Данилов, 1975). Неблагоприятными оказываются также зимы с очень низкими температурами при отсутствии снежного покрова достаточной толщины, а также зимы с частыми сменами морозов, и оттепелей. Высокая численность птиц осенью может стать причиной опустошительных эпизоотий, наблюдавшихся в Уссурийском крае (Воробьев, 1954) и на Урале (Данилов, 1975), а также увеличивает отрицательное воздействие хищников и, наконец, вызывает усиление дисперсии особей из перенаселенных мест.
Среди хищников, влияющих на численность рябчика, необходимо упомянуть ястребов (тетеревятника и перепелятника), лисицу, росомаху, куницу, соболя, горностая. Рябчик может стать добычей практически всех крупных хищников, обитающих в данной местности, и общий список последних, в рационе которых он отмечен, довольно велик. Вполне очевидно, что редкие виды, как беркут или орлан белохвост, не могут заметно влиять на его численность. Наиболее же заметно сказывается деятельность куньих. Например, в верховьях р. Селемджи зимой 80% гибели птиц приходится на соболя — один зверек за зимний сезон поедает до 25 рябчиков (Юдаков, 1968). В других же районах в отдельные годы процент встречаемости рябчика в экскрементах этого хищника не превышает 1.7 (Прибайкалье; Измайлов, Павлов, 1975). Значительна доля рябчика и в рационе куницы. В верховьях Западной Двины, например, неоднократно находили даже запасы, сделанные этим хищником из добытых им рябчиков. Такие «склады» обычно помещались в дуплах осин и состояли из целых или слегка поеденных упитанных птиц (Волков, 1975).
Неурожай какого-либо из основных кормов тоже может повлиять на численность птиц, но сравнительно высокая эврифагия рябчика заметно смягчает отрицательное воздействие таких явлений.
Высокая численность рябчиков, достигаемая благодаря сочетанию благоприятных обстоятельств, под воздействием процессов дисперсии, деятельности хищников и влияния заболеваний сохраняется не более 1—2 лет, после чего она несколько лет равномерно понижается (Романов, 1975). Бывает и так, что численность рябчиков резко падает в первую же зиму после вспышки.
Наиболее оптимальным для гнездования рябчика являются центральные части ареала в подзоне южной тайги, имеющие умеренно континентальный климат. Судя по многим данным, нормальной следует считать плотность, которая достигает весной 10—15 пар на 1 км2, а осенью — 30—50 птиц на ту же площадь. При низкой численности весной одна пара приходится нередко на 5—6 км2 подходящих местообитаний.
Популяционный состав (численное отношение полов).
Соотношение полов у взрослых птиц близко 1:1. У сеголетков бывает значительное (на 12.5 %) преобладание самцов (Гайдар, 1974). Убыль молодых птиц к сентябрю составляет 24—50 % (Донауров, 1947; Данилов, 1975; Дулькейт, 1975), хотя, вероятно, метод подсчета убыли молодых птиц по количеству их в выводках может сильно завышать действительные цифры: в августе уже происходит распад выводков.
Территориальность.
Рябчик — одна из наиболее оседлых тетеревиных птиц. Оседлый образ жизни обусловлен обилием корма практически во все сезоны года, что делает ненужным сезонные перекочевки. Смена сезонных кормов сопровождается только небольшими перемещениями из одних типов леса в другие. Максимальное удаление от места кольцевания — 5 км (Скандинавия, бассейн Печоры; Семенов-Тян-Шанский, 1959; Гайдар, 1973). 90 % окольцованных птиц не удалялось от места кольцевания далее 500 м, причем наиболее далеко перемещались старые самки и молодые птицы обоих полов. По данным Романова (1975), из 37 окольцованных птиц 34 были встречены не далее 700 м от места выпуска и только 3 — на расстоянии 1000—2000 м. Им же отмечен случай, когда один и тот же самец 3 года держался на одном участке. В период размножения территориальная структура популяции представлена системой участков, то распределенных сравнительно равномерно, то группирующихся по 2—4 в особо удобных местах. Эти участки — гнездовые, где птицы! обычно обитают парами. Связь с территорией особенно сильна в период размножения. У западных популяций она сохраняется и зимой, тогда как у восточных популяций, в особенности восточнее Енисея, индивидуальных территорий в зимнее время обычно нет и птицы ведут стайный, хотя и вполне оседлый образ жизни. В долине р. Омолон, например, участок обитания зимней стаи постоянен, его размеры — около 1 км2, причем в середине зимы размеры его сокращаются до 0.5 км2 (Андреев, 1980).
Система гнездовых участков нарушается со второй половины лета. Выводки нередко выходят за их пределы, перемещаются в иные биотопы (например, на ягодники), нередко объединяются в крупные, хотя и временные скопления, и так до второй половины осени, когда вновь, по крайней мере у западных популяций, происходит распределение территории между отдельными птицами, в том числе и молодыми. Индикатором этого процесса является осеннее токование. Остается, однако, невыясненным, насколько успешно молодые птицы в это время занимают участки. Что же касается образования пар, то, по одним данным, они образуются осенью редко, а возникнув, распадаются к весне (Punnonen, 1954), по другим (Гайдар, 1974) — это обычное явление, причем в формируёмых осенью парах нередко одна из птиц — молодая. Есть данные, что осенью ведущее значение в выборе и закреплении участка играет самка — свист ее в это время слышен гораздо чаще, чем весной (Риппбпеп, 1954). По моим наблюдениям, основным связующим звеном для пары является гнездовая территория. При гибели одного из партнеров его заменяет другая птица того же пола, но территория остается прежней. Например, в каком-нибудь лесном массиве имеется ряд участков, где постоянно, из года в год можно встретить рябчиков, в то время как в прилегающих местностях эти птицы водятся далеко нерегулярно.
Образ жизни.
Проводя всю жизнь под пологом леса и избегая даже при местных перемещениях подниматься выше верхушек деревьев, рябчик, видимо, вообще не способен к длительным перелетам через открытые пространства. Только в редких случаях; он отваживается перелетать долины рек или горные ущелья, не более 300—500 м над открытой местностью, и, стремясь при этом как можно скорее вернуться под покров леса. Полет этой птицы в лесу отличается высокой маневренностью. Набрав скорость дюжиной быстрых энергичных взмахов, он планирует затем на расставленных крыльях и, меняя их положение, ловко маневрирует среди густой растительности.
Большую часть времени птицы проводят на земле, где в теплое время года добывают корм, гнездятся, водят выводки, совершают ряд процедур по уходу за оперением, а иногда и спят. На деревьях кормятся преимущественно в холодное время года, но и зимой большую часть времени проводят на снегу (в подснежной камере). Рябчик легко и свободно перемещается по самому захламленному субстрату и при необходимости может быстро бегать. В случае опасности далеко не всегда взлетает, уходя нередко пешком. Осенью и весной, а также зимой во время оттепелей птицы, как правило, ночуют на деревьях, забившись в гущу еловых или пихтовых ветвей. На деревьях рябчик также чувствует себя вполне свободно и, добираясь до концевых сережек, может передвигаться по самым тонким веточкам.
Будучи внезапно испуганным, рябчик взлетает на одно из ближайших деревьев и некоторое время осматривается, выясняя степень опасности. При наличии последней он перелетает еще несколько десятков метров и садится так, чтобы быть невидимым. Затаившись, он сидит совершенно неподвижно и иногда не взлетает даже при выстреле — это особенно характерно для птиц, впервые сталкивающихся с человеком. Вообще же поведение рябчиков в различных местах и при различных условиях может сильно меняться. Так, например, в сплошных березовых лесах Речицкого Полесья после опадания листьев вспугнутые птицы не садятся на голые березы, а летят как можно дальше и садятся вновь на землю, как фазаны, стараясь укрыться в зарослях или валежнике (Федюшин, 1959). В ненаселенных же местах Приполярного Урала, по нашим наблюдениям, рябчики, будучи вспугнутыми, часто садились на березы, совершенно не таясь.
Рябчик — довольно молчаливая птица. Наиболее обычный позыв самца — свист, который особенно громко и часто звучит во время токования, но может быть услышан в любое время года. Самки тоже свистят, хотя свист их короче, проще и тише. Тон и ритмика свиста как у самцов, так и у самок могут существенно варьировать. Основная и явно первоначальная роль такого свиста — установление контакта между особями, а во время токования он же выражает степень полового возбуждения птицы и выполняет роль брачной песни. Более разнообразны голосовые реакции, предупреждающие выводок об опасности (см. далее). При сильном волнении или испуге как самцы, так и самки издают гортанный вскрик, наподобие испуганной курицы. К тревожным позывкам относятся также и своеобразная звонкая трелька, которую можно слышать от вспугнутых птиц — она звучит как следующий друг за другом с убыстрением ряд отрывистых звуков «пить... пить... пить... пить-пильпильпиль».
Повадки и образ жизни в весенне-летний период.
В весенне-летнее время рябчикам свойствен ряд специфических поведенческих реакций, связанных с уходом за оперением и очищением его от эктопаразитов. Это так называемое муравьение и пылевые (песочные) ванны. При «муравьении» рябчики забираются на только что оттаявшие муравейники, на макушках которых начинают копошиться муравьи, и, топорща перья, провоцируют нападение муравьев, обрызгивающих перья птиц едким секретом, так называемой муравьиной кислотой. Этот секрет освобождает птиц от перьевых клещей. «Муравьение» наблюдается только ранней весной, когда окончившие спячку муравьи еще не кусаются, но щедро выделяют этот секрет (Дубинин, 1956; Формозов, 1976). Пылевые ванны принимаются птицами на специальных сухих местах, хорошо прогреваемых солнцем, где птицы либо выбивают сами, либо используют естественные ямки в сухом пылеватом субстрате диаметром 13—15 см и глубиной около 6 см, называемые «порхалищами» (Фетисов, 1976).
Здесь птицы регулярно, с начала мая по конец августа, проводят около получаса в день, «купаясь» в пыли или песке, забрасывая его на спину, всячески стремясь забить побольше сухого пылевидного субстрата между перьями. Вслед за этим птицы встряхиваются, по взъерошенным перьям пробегает вибрирующая дрожь и набившийся туда субстрат при этом выпадает, увлекая за собой эктопаразитов. Обычно на гнездовом участке имеется 1, реже 2—3 таких места (Klaus et al., 1976).
Образ жизни в зимний сезон.
Весь зимний сезон рябчики остаются строго оседлыми. В европейской части СССР (Калининская и Ленинградская области) зимний участок рябчика занимает от 2 до 4 га, причем расстояние между участками может достигать 120—580 м. Только в наиболее благоприятных местах они могут примыкать друг к другу (Волков, 1968в; данные автора). Все зимнее время рябчики западных популяций держатся парами или одиночками, иногда собираясь на кормных местах по 4—6 птиц. Так, в Калининской обл. 55 % рябчиков зимует поодиночке, 42 % — парами и 3 % — группами по 3 птицы (Волков, 1968в). На востоке ареала эти птицы собираются зимой в довольно большие стаи, по 15—25 особей, причем состав этих стай довольно постоянен. Во всяком случае на Омолоне, в приохотской тайге, в Приморье, на Сахалине и на Хоккайдо рябчики не только кормятся стайками, но вместе же и ночуют (Андреев, устное сообщ.; Гассовский, 1927; Taka-Tsukasa, 1932; Сингиревский, 1946; Воронов и др., 1975). В ряде мест (Северный Урал, Алтай) рябчики нередко кормятся стайками по 8—12 птиц, но совместных ночевок при этом не наблюдалось (Донауров, 1947).
В течение всей зимы рябчики кормятся преимущественно (а в сильные морозы — исключительно) на деревьях. При слабых же морозах или в оттепели они нередко бродят по снегу, проходя от 100 до 300 м за одну кормежку, поедая почки и верхушечные части побегов подроста берез, осин, ольхи, ивы, кустиков малины и черной смородины, торчащей из-под снега черники. Это особенно характерно при наличии обильного снега на ветвях (кухты), причем в таких ситуациях снег нередко пригибает к земле верхушки молодых берез и ольхи, которые могут даже вмерзать в снег. Эти вершины нередко несут на себе значительное количество сережек и почек, и птицы кормятся ими, расхаживая вокруг по снегу. Скорость передвижения рябчика при пешей кормежке — около 2 м в минуту.
В условиях морозной и снежной зимы рябчик большую часть суток проводит в подснежной камере. По моим наблюдениям, птицы предпочитают зарываться в снег не с лёта, а предварительно сев на него и пройдя несколько шагов.
В тоннеле рябчик нередко делает 1—3 контрольных отверстия, высовывая голову на поверхность. Нередко, пройдя около 1 м птица делает резкий поворот на 90° и прорывает тоннель дальше еще на 0.4—1 м, а затем отрывает камеру, которая имеет типичное для всех тетеревиных птиц устройство. На все время отрывания камеры вместе с тоннелем уходит 20 с (Андреев, 1980). Размеры камеры: ширина 10—11 см, длина 18—19 см, высота 12—15 см. Толщина потолка колеблется в зависимости от толщины и состояния снега от 1 до 10 см. В среднем расстояние от пола камеры до поверхности снега составляет 21—23 см. При наличии наста птица ищет места, где он менее прочен, обычно в зарослях кустарников. Если же крепость наста не позволяет погрузиться под снег, рябчики ищут хотя бы какое-нибудь подобие лунки, естественную ямку, которую стараются по возможности углубить. Ежедневное копание в снегу, проделывание тоннелей и отрывание камер приводят к сильной изношенности оперения верхней части тела, что особенно хорошо заметно на коллекционных экземплярах из восточной части ареала. В дневное время, если мороз не очень силен, рябчики отдыхают на солнцепеке в открытой лунке глубиной в 10—16 см. Рябчики могут устраивать подснежные камеры только тогда, когда глубина снега превысит 20 см. Во время морозов при малоснежье птицы ищут малейшей возможности зарыться в снег, хотя бы частично. В таких случаях птица зарывается наискосок к плоскости грунта и снаружи может торчать хвост (Андреев, 1980). При атмосферных температурах выше —10 °С в условиях малоснежья рябчики могут ночевать и на деревьях (в оттепели — как правило), забиваясь в густые ветви елей или пихт. В малоснежных районах с морозной зимой (Якутия) рябчики накапливают к зиме значительные жировые отложения (Егоров и др., 1959). Вообще же в северных районах эти птицы, как и некоторые другие тетеревиные, накапливают осенью жир даже при регулярно образующемся там глубоком снежном покрове. Например, на Омолоне (65° с. ш.) рябчики накапливают значительные жировые запасы уже в октябре, причем в отдельных случаях количество жира может составлять 11 % от массы птицы, но обычно его бывает меньше, около 4 % (Андреев, 1980). На Приполярном Урале (66° с. ш.), по моим наблюдениям в 1974 г., рябчики стали накапливать жир с начала октября, еще до выпадения снега, при положительных температурах. С установлением в середине октября зимней погоды они перешли на питание преимущественно сережками березы и буквально за неделю, к 22—23 октября, стали очень жирными. Жир откладывался на кишечнике, вокруг основания хвоста, у основания шеи и на бедрах. Птицы начали ночевать в снегу с 20 октября, когда глубина его достигла 40 см, а температура воздуха ночью стала падать до —15 °С. Продолжительность ночи в это время составляла уже 16 ч.
Ежесуточная жизнь рябчика зимой складывается из следующих типов деятельности: пребывания в подснежной камере в состоянии покоя, передвижения по ветвям деревьев или по снегу при поиске и добывании корма и самой кормежки, отдыха в середине дня на поверхности снега или на дереве и, наконец, перелетов. Затраты времени на все эти виды активности зависят прежде всего от атмосферной температуры и характера погоды, а также от длины ночи, от наличия тех или иных кормов, от состояния снежного покрова и степени заснеженности деревьев. В середине зимы в северной части ареала при сильных морозах (ниже —20 °С) рябчики проводят в подснежных камерах почти все сутки (около 23 ч), затрачивая на кормежку немногим более часа. В наиболее суровых условиях в самое темное время (66° с. ш., декабрь, —40 °С) рябчики кормились дважды — утром и вечером — по 30—40 мин, затрачивая суммарно на кормежку 1.2 ч. 3—3.5 ч дневного времени между кормежками проводились также в подснежной камере. Там же, при среднесуточных температурах —30 °С, птицы имели только одну, утреннюю кормежку, но длительностью в два раза большую (Андреев, 1980). На 60° с. ш. (Ленинградская обл.) при умеренных морозах —10-20 °С в условиях солнечной погоды происходит либо одна утренняя кормежка длительностью от 1.5 до 4 ч с пребыванием в остальное время в подснежной камере, либо 2-часовая утренняя кормежка, затем отдых на солнцепеке, обычно в поверхностной, открытой лунке в течение 2—4 ч, после чего вновь короткая кормежка и уход на ночлег. Чем теплее погода, чем тверже поверхность снега, чем длиннее день, тем более длинной и менее интенсивной становится кормежка, тем чаще она прерывается паузами отдыха по 40—70 мин каждая. При небольшом насте в середине зимы во время умеренных морозов (—15 °С) некоторые особи могут в середине ночи 1—2 раза менять место ночлега, проходя по 5—10 м в поисках более удобного места и вырывая новую камеру. Будучи потревожен в ночное время хищником, рябчик без труда может взлететь и, пролетев несколько десятков метров, вновь сесть на снег и зарыться в него.
Кормовая активность, рацион, его состав и количество довольно сильно зависят от температурных и погодных условий. В экстремально суровых ситуациях (как, например, на Омолоне) птицы, резко сокращая время кормежки, поедают наиболее легкодоступный, хотя и менее питательный корм (почки ив, побеги берез, ивы и чозении), причем общее его количество при этом увеличивается. При скусывании веточек и почек в таких условиях у птицы, кормящейся на ветвях, интервал между поеданием отдельных кусочков составляет 1.38—1.47 с, тогда как на отыскание и поедание одной сережки уходит от 17 до 50 с (Андреев, 1980). Чем теплее погода и чем длиннее день, тем больше энергии и времени может затрачивать птица на поиск корма и доля сережек в рационе пропорционально возрастает. При температуре, близкой к 0 °С, рябчики нередко кормятся, расхаживая по поверхности снега и поедая почки с подроста и кустарника. Ходьба по рыхлому снегу для него, видимо, затруднена — проваливание может достигать 5 см.
Самая низкая температура, при которой была отмечена кормежка рябчика, —49 °С. В этом случае птицы кормились только 25 мин. Оперение их было распушено, движения замедлены, причем птицы замирали время от времени на 3— 5 мин, видимо, для того, чтобы корм в зобу успевал прогреваться (Андреев, 1980).
При низких температурах рябчики затрачивают на полетную активность самое минимальное время — не более 2 мин. 1 или 2 раза в сутки птицы делают короткий, не более 100 м перелет на кормовые деревья и обратно, с места кормежки к месту ночлега — примерно на такое же расстояние.
Обращает особое внимание стремление птиц в условиях крайне низких температур использовать все возможные способы для уменьшения теплопотери. При солнечной погоде даже в сильные морозы они могут, сильно распушив оперение, греться в косых лучах солнца, сидя на верхушках деревьев, а при ночевках выбирают для устройства подснежных камер самые низкие места поймы, где наиболее ощутим тепловой поток, идущий от земли (Снигиревский, 1946; Андреев, 1980). Сообщения о том, что во время метелей, а также при морозах ниже —35 °С рябчики могут проводить в подснежных камерах двое и более суток подряд (Снигиревский, 1946; Дулькейт, 1964), не подтверждены.
Величина суточного рациона птиц в зимнее время явно меняется, но в небольших пределах, будучи зависимой и от характера корма, и от физиологического состояния птицы, и от погоды. При низких температурах и коротком световом дне рябчики уходят на ночевку с максимально наполненным зобом, который содержит количество корма, достаточное для поддержания существования птицы до следующей утренней кормежки. Максимальное количество корма отмечено в зобах двух рябчиков, добытых на Омолоне в декабре при температуре ниже —40 °С, 81.2 и 84.6 г сырой массы. Обычно же вечером максимальное содержимое зоба составляет: 59.5, 56 г (Омолон, Андреев, 1980); 47, 48, 54, 55, 58 г (Семенов-Тян-Шанский, 1959; Кольский п-ов); 55, 58 г (Приполярный Урал, данные автора). Вычисленный по этим данным суточный рацион составляет 81.3 г сырой массы (сухая масса— 47.97 г). Его калорийность составляет в среднем 1068 кДж.
Размеры твердых экскрементов рябчика зависят от пола и возраста птицы и колеблются в пределах: 17.2—24x5—7.3 мм, масса — в среднем 0.159 г (0.116—0.178). Общее их количество в ночных камерах колеблется от 60 до 85. Для вычисления энергии, удаляемой с экскрементами, количество ночевочных экскрементов экстраполируется на все сутки, что может дать заниженный результат. По моим данным, корм из зоба может быть израсходован раньше утреннего выхода на кормежку и какое-то время перед выходом из камеры птица вообще не выделяет экскрементов. Кроме того, время от начала кормежки и до появления первых экскрементов может составить у рябчиков до 1.5 ч (Bump et al., 1947). И, наконец, в дневное время скорость выделения твердых экскрементов, видимо, возрастает, составляя 7 мин на один экскремент (Е. Р. Потапов, устное сообщ.) против 12 мин в ночное время. С учетом всех этих моментов мои расчеты дают 123 экскремента в среднем на птицу в сутки, т. е. получается примерно та же величина, если бы мы исходили из равномерности темпов пищеварения в течение всех суток и непрерывности выделения экскрементов со скоростью 12 мин каждый.
Экскременты слепого отдела кишечника (выброс) выделяются птицей, как и у остальных тетеревиных, обычно при оставлении ночной камеры. Однако, покидая ее, рябчик может оставить выброс не сразу, а спустя некоторое время, когда уже начал кормиться на деревьях на значительном удалении от ночевки. Мне неоднократно приходилось находить выбросы там, где в радиусе 50 м не было никаких признаков ночевки птицы. Кроме того, поскольку экскременты из слепых кишок выделяются в большинстве случаев 3—5 порциями, у ночевочной камеры может остаться только часть выброса, а остальная будет выделена в другом месте. Поэтому при расчетах нужно использовать только те случаи, когда нет никаких сомнений, что весь выброс находится у камеры или что его действительно не было до начала кормежки птицы. Последнее бывает возможно установить или прямым наблюдением или только тогда, когда птица, покидая ночевку, не взлетает, а проходит некоторое расстояние и начинает кормиться пешком. По моим данным, сухая масса выброса колеблется в пределах 3—8.65 г, составляя в среднем 5.13 г.
Калорийность твердых экскрементов составляет у рябчика около 21.55 кДж/г, а калорийность выброса 26.5 кДж/г (Андреев, 1980). Рассчитанная на основании этих данных энергия существования в зимнее время составляет 400 кДж/особь/сутки для птиц Ленинградской обл. при температуре —20 °С и 422.88—481.5 кДж/особь/сутки для Омолона при температуре —40 °С (Андреев, 1980). Данные по Омолону, характеризующие энергию существования рябчика в два разных зимних сезона, показывают, что величина энергии существования может заметно меняться в зависимости от условий зимы даже в одной и той же местности.
Питание.
Основу зимнего рациона составляют сережки, почки и концевые побеги березы, ольхи, ивы и тополя. Чаще всего поедаются сережки березы или ольхи — в зависимости от степени обилия тех или иных сережек и их доступности. При малом количестве сережек, равно как и при сильных морозах, увеличивается доля почек и концевых побегов. В Сибири у добытых птиц в зобах насчитывалось до 2000 кусочков веток и почек березы (Дулькейт, 1960). Поедая концевые побеги ольхи, рябчик может откусывать кусочки до 2 см длиной, при диаметре 2—2.5 мм. Древесина побегов нередко составляет основную массу в желудке, задерживаясь здесь дольше всего. При оттепелях птицы поедают в небольших количествах стебли черники, скусывая верхушечные побеги. Зимнее питание рябчика по всему ареалу не столь однообразно, как принято думать. Оно существенно меняется с запада на восток — береза и ольха уступают место тополю (чозении) и иве. Удельный вес веточного корма начинает заметно падать с момента установления положительных температур и постепенно сходит на нет к концу весны, хотя в малом количестве этот корм присутствует все лето. С исчезновением снежного покрова рябчик постепенно переходит на наземное питание, копаясь в сырой подстилке и поедая зеленые побеги, почки и перезимовавшие ягоды, семена и плоды (в том числе и семена ели), цветки, побеги, листья (особенно цветки и побеги ветреницы), а позже и семена различных трав, иногда зеленую хвою пихты. Однако всю весну нередко преобладают древесные корма, например, почки и листья рябины, почки осины составляя до 66 % содержимого зобов (Олигер, 1973).
Летнее питание довольно разнообразно, особенно у южных популяций. Качественное отличие летне-осеннего питания (июнь—сентябрь) — значительная доля животных кормов. Поедаются большей частью различные насекомые (жуки, муравьи, кузнечики, разнообразные гусеницы и личинки), пауки, моллюски (в основном слизни). Но и в летние месяцы доля животных кормов не превышает 5 % рациона (Олигер, 1973). Примерно с июля, а в южных частях ареала с середины июня в рационе появляются различные ягоды, которые в конце лета и осенью, вплоть до октября становятся основным кормом. В северных частях ареала главное место в питании занимают черника, брусника и голубика, причем ягоды начинают поедаться еще зелеными. В южных частях ареала состав ягодного рациона значительно разнообразнее. Вообще рябчики поедают практически любые ягоды, и ягодный рацион зависит скорее всего от видов и урожая встречающихся в данной местности ягодных растений.
Переход на зимнее питание начинается в сентябре, когда птицы уже регулярно поедают почки и сережки березы, ольхи, ивы, рябины, осины и других лиственных деревьев. Чем ниже температура воздуха, чем длиннее становится ночь, тем большее место занимают они в питании и становятся основным, а нередко и единственным кормом еще до образования сплошного снежного покрова. В отдельных случаях, например при большом неурожае ягод, переход на зимнее питание может совершаться еще раньше. Так, например, в южном Приангарье после большого неурожая ягод в 1971 г. (все уродившиеся ягоды были полностью использованы птицами к концу августа), рябчики перешли на зимний рацион на месяц раньше обычного (Владышевский, 1975б).
В восточных частях ареала в зависимости от наличия и урожая кедровой сосны большое значение в осеннем питании может иметь кедровый орех, который либо собирается с земли, либо даже выклевывается из шишек. Заглатываются орешки целиком и размельчаются уже в желудке. В различных частях ареала существенную часть осеннего рациона могут составлять ягоды рябины и семена ели, хотя при урожае последней семена вылетают из шишек осенью очень редко. Основная масса семян выпадает с начала марта до конца мая, и рябчики в это-время охотно питаются ими, расхаживая по снегу. При обилии семян рябчик даже во время интенсивного токования может приобретать на такой диете заметные жировые отложения (Формозов, 1976). На Омолоне отмечено осеннее питание семенами лиственницы в урожайные годы — находили до 2500 семян в одном зобу, причем птицы на этом корме также быстро жирели (Андреев, 1980).
В питании птенцов рябчика заметную долю составляют животные корма. Первые полторы-две недели птенцы питаются исключительно животной пищей — мелкими пауками, личинками пилильщиков, цикад и комаров, муравьями, гусеницами пядениц и совок и т. п. Далее рацион разнообразится мелкими семенами различных трав (кислица, лютик, фиалка), хотя основу питания по-прежнему составляют пауки и насекомые. По мере созревания ягод птенцы переходят на ягодный рацион, но вплоть до конца августа—начала сентября процент животного корма в рационе молодых птиц по сравнению со взрослыми остается значительным — до 20. Только со второй половины сентября разница в питании молодых и старых птиц исчезает. Количество гастролитов в желудке (камешки, косточки ягод) колеблется в зависимости от сезона, хотя строгой закономерности здесь нет. Камешков меньше всего в августе, когда при ягодном рационе роль гастролитов выполняют, хотя по мнению некоторых и малоуспешно (Фолитарек, 1938), твердые семена и косточки плодов и ягод. С августа количество камешков в желудке быстро увеличивается, достигая максимума в ноябре. Бывает, однако, что в некоторых случаях максимальное количество камешков в желудках самок (до 8.1 г) наблюдается весной (Ленинградская обл.; Родионов, 1963б), хотя обычно количество гастролитов к весне падает.
Размножение и токование.
Рябчик — моногам, и по крайней мере в западной части ареала птицы большую часть года живут парами. Степень постоянства пар не выяснена, хотя известны случаи, когда одни и те же птицы держались в паре два года подряд. Характер токования самца — одиночный. Весеннее оживление наступает в начале марта на юге ареала и в середине или в конце месяца — на севере. Самцы начинают расхаживать по снегу со слегка расставленными и приспущенными крыльями, концы которых волочатся по снегу, оставляя по краям следов характерные бороздки, «чертежи». При встрече друг с другом самцы иногда начинают двигаться по снегу параллельно на расстоянии 1—2 м друг от друга с приспущенными крыльями и пробегают при этом несколько десятков метров. Цель такой «параллельной пробежки», неясна, во всяком случае она явно не имеет отношения к охране гнездового участка. В целом же токование на первых порах протекает вяло, самцы свистят и бегают по снегу в токовой позе только в хорошую погоду и, как правило, только по утрам. У птиц, которые зимуют стайками, весеннее возбуждение может спонтанно возникать и в стае. Случаи такого рода наблюдались в Печоро-Илычском заповеднике в стаях рябчиков по 15 и но 17 особей. В обоих случаях вся поляна, на которой находились птицы, была испещрена следами самцов, бегавших с распущенными и волочащимися по снегу крыльями (Донауров 1947).
Брачная активность птиц достигает максимума в то время, когда в лесах средней полосы дотаивают последние пятна снега. По мере его стаивания самцы все более активно защищают свои участки, и в самый разгар токования (середина апреля — для южной части ареала, конец апреля—начало мая — для средней и конец мая — для северной) между соседними самцами все время происходят стычки, причем вторгающийся на чужую территорию самец, как правило, изгоняется. Самка обычно находится неподалеку от самца и большую часть времени занята кормежкой. Временами она отвечает на свист самца и внимательно следит за стычками, в которых он участвует. На чужую самку, появляющуюся в пределах участка, самец не нападает, но следит за ней до тех пор, пока она не покинет участка (Ивантер, 1962). В разгар токования происходит и спаривание.
Рябчики достигают зрелости на первом же году жизни. Начало развития гонад у самцов не зависит от погоды, находясь целиком под фотопериодическим контролем. Так, в Ленинградской обл. оно начинается после того, как длительность светового дня достигает 13 ч (Родионов, 1963б). Сроки же максимального развития семенников зависят от хода весны и могут колебаться в пределах 30 дней. Степень развития гонад у самок в еще большей степени зависит от погодных условий и прежде всего от появления достаточно больших участков свободной от снега земли — именно в это время и начинается спаривание. Однако при слишком раннем стаивании снега (ранняя и теплая весна после малоснежной зимы) самки не провоцируются этим и начинают кладку примерно в те же сроки, что и обычно. Известны, правда, отдельные случаи, когда в очень теплые ранние вёсны самки могут начинать кладку на 2 недели ранее обычного (Ленинградская обл., сообщения егерей). В течение всего периода интенсивного токования самец очень активен. Сопровождая кормящуюся самку, он то и дело издаёт характерный свист, совершает ритуалы ухаживания (см. далее) и демонстрационные полеты, а в состоянии особого возбуждения — токовые взлеты и бурно реагирует на вторжения в пределы, его участка посторонних самцов. В такой ситуации самец-хозяин принимает сначала настороженную позу (рис. 9, 2), при которой хохол поднят, а белое пятно на шее максимально расширено, затем он свистит один или несколько раз и, получив ответ, устремляется к противнику по воздуху или по земле. Во время конфронтаций дело редко доходит до драки, поскольку, как правило, посторонний самец покидает участок при первом же натиске хозяина.
Брачный ритуал самца состоит из песни, токового полета, токового взлета и особых поз, сопровождающихся специфичными вокальными сигналами. Песня рябчика — совершенно особая и, за исключением B. sewerzowi, не имеет аналогов в пределах семейства. Это протяжный тонкий свист, длящийся около 3 с и состоящий из прерывистых сигналов — двух длинных (второй несколько короче первого) и последующих коротких, которые, быстро следуя один за другим, сливаются в прерывистую трельку. Частота свиста — около 7000 кГц (Hjorth, 1970). Слышимость его при хороших условиях — около 100 м. Обычно свист каждого самца имеет свои индивидуальные особенности (в основном в ритмике), но и каждый самец может давать различные вариации (Bergmann et al., 1975). Фонограммы свиста самца из различных мест ареала показывают существенные различия в манере исполнения (рис. 8), которые в некоторых случаях (Франция) имеют явно популяционный характер. По некоторым данным (Fuschlberger, 1956), свист рябчика сопровождается резким шелестящим звуком, слышным только на самом близком расстоянии. На сонограммах следы этого звука не прослеживаются.
Свистящий рябчик сидит на ветке или стоит неподвижно на земле, глубоко втянув шею и открыв клюв (рис. 9, 2), хотя до сих пор бытует мнение, будто рябчик свистит, вытянув шею. Примечательно, что улары свистят, также широко открывая клюв. Во время свиста самец слегка распускает хвост, крылья прижимает к телу, лишь слегка отставляя их кончики. Клюв широко открывается с каждым звуком, и хвост синхронно вздрагивает с ритмом свиста (Punnonen, 1954). Во время свиста рябчик несколько снижает свою бдительность (Scherzinger, 1978). Свист самки короче и состоит из 3 нот, первая и последняя из которых примерно такой же тональности и длительности, как первая часть песни самца, вторая же несколько длиннее и иногда становится как бы двусложной.
Токовой полет самца выглядит вначале как обычное перемещение с дерева на дерево. Но заканчивая его, птица замедляет темп взмахов, и крылья продуцируют особенно громкие хлопки. Хвост при этом раскрыт на 125°. Токовые взлеты не занимают особо важного места в ритуале, но они очень интересны, поскольку являютсй как бы первой стадией того развития, которое привело к выработке «барабанной трели» B. umbellus. У B. bonasia они исполняются по-разному у разных подвидов. Так, в Финляндии самец взлетает вверх на 1—2 м, приземляясь на землю примерно в том же месте. Крылья при этом работают с громким быстрым хлопаньем. Иногда самцы производят серии таких хлопков, но не взлетая, а только слегка приподнимаясь и придавая телу более вертикальное положение (Punnonen, 1954). В Западной Европе, наоборот, самцы взлетают редко и, как воротничковый рябчик, как бы обозначают такой взлет на месте, приподнимаясь вертикально и энергично работая крыльями. В некоторых случаях хлопки крыльев следуют друг за другом с такой частотой и громкостью, что могут быть приняты за барабанную трель дятла (Teidoff, 1952; Fuschlberger, 1956). Эти хлопки слышны примерно за 100 м. Выполняя такой взлет в вольере, рябчик поднимается на 0.5—1.5 м, но пролетает при этом 2—4 м (Scherzinger, 1978).
У восточных популяций существует еще несколько элементов брачного поведения, пока не обнаруженных у западных. Это прежде всего «сигнал места», как назвал его А. В. Андреев, — громкое шипение, издаваемое самцом в определенной позе (рис. 9, 3) после завершения токового полета (подвид B. B. kolymensis; Кречмар и др., 1978). Это шипение описано затем и для западноевропейских птиц, содержавшихся в неволе, — шипящий звук, как у тетерева, только в отличие от восточных птиц очень тихий издавался птицами в вертикальной позе, обычно в связи с токовым взлетом (Scherzinger, 1978). У самцов восточных популяций я наблюдал также своеобразное урчание, издаваемое при наличии поблизости соперника. Это урчание представляет собой несколько следующих друг за другом одинаковых фраз, каждая из которых начинается отрывистыми слогами «вить-вить-вить...». Эти звуки быстро учащаются, сливаясь в короткую булькающую трельку, заканчивающуюся свистом, который резко меняет свою тональность с высокой на низкую. После этого свиста вновь следует завершающие булькающие звуки с понижением тона в конце. Длительность такого урчания — около 4 ч, она несколько напоминает аналогичную голосовую реакцию гималайского улара Tetraogallus himalayensis Gray. Это урчание-самец издает в моменты сильного возбуждения — «брови» максимально расширены и заметно выступают над теменем, шея изогнута и выглядит сильно расширенной, хотя перья на ней не топорщатся. Мне нередко приходилось слышать это урчание в разгар токования на Нижнем Амуре и ни разу — у рябчиков в европейской части СССР. Не упоминает о таком урчании и Пюннонен, детально описывавший голосовые реакции рябчиков. Однако недавно нечто подобное было описано для западноевропейских рябчиков, содержавшихся в вольерах (Scherzinger, 1978). Урчание свойственно и рябчикам Забайкалья (С. П. Кирпичев, устное сообщ.).
Основная токовая поза у самца такая же, как и у большинства лесных тетеревиных птиц. Полностью раскрытый хвост поднят почти вертикально, как и слегка вытянутая шея, крылья несколько отставлены в стороны и приспущены. Перья на груди, боках и брюшке приподняты, что создает эффект увеличения размеров птицы. На шее же перья поднимаются почти вертикально, резко контрастируя с гладким оперением головы, кажущейся при этом несообразно маленькой.
В агрессивной позе самец также поднимает перья, немного поднимает полураскрытый хвост, вытягивает вперед шею и голову и, издавая короткие резкие звуки, бежит навстречу сопернику. При этом перья на подбородке стоят торчком, образуя что-то похожее на «бороду» самца глухаря (рис. 9, 4).
При столкновениях самцы стоят друг перед другом в вертикальной позе, с удивительной синхронностью совершая взаимные угрожающие выпады, — каждому выпаду одного из них точно соответствует обратное движение соперника. Удары наносятся клювом в голову и шею, а также обоими крыльями и ногами. Удары ног бывают наиболее сильными и наносятся во время прыжков (Scherzinger, 1978). Последний автор описал еще очень важную деталь, не наблюдавшуюся пока в природе, — активное и громкое топтание самца на месте.
Иногда самец, в особенности при виде пролетающей поблизости самки, издает призывной свист, напоминающий позывку летящего лугового конька. Свист этот очень изменчив и буквами его можно передать, как «ит-иррррр» (Punnonen, 1954).
В предкопуляционном поведении выделяются два ритуальных компонента. Сначала самец кружит вокруг самки в основной токовой позе, раскрытый хвост слегка повернут в ее сторону, а голова, наоборот, в противоположную сторону (рис. 9, 5). Затем, непосредственно перед спариванием оба партнера, располагаясь спиной друг к другу, прижимаются к земле и начинают резко поворачивать головы синхронно, но в разные стороны (Scherzinger, 1978). Эта процедура завершается спариванием, во время которого самец удерживает самку за хохол (Hoglund, 1957). Согласно другому описанию (Lindemann, 1952), перед спариванием самец, приближаясь к самке, садится к ней почти вплотную. Возбуждение его проявляется в усиленном свисте, тело наклонено вперед, голова с торчащим хохолком и открытым клювом также вытянута вперед, перья на подбородке и шее топорщатся. Возбужденное дрожание и подергивание тела становится все сильнее, при этом крылья бывают раскрыты, а хвост подергивается вниз. Глаза то и дело затягивает мигательная перепонка, свист продолжает звучать в полную силу. Самка принимает приглашающую позу, и самец «топчет» ее с распущенными крыльями и хвостом, с поднятым хохолком и максимально расширенными «бровями». Спаривание происходит только на земле.
Одиночный характер токования — правило, но в местах с высокой численностью этого вида (Сибирь, Дальний Восток) во время токования в отдельных местах можно встретить вместе до 4 самцов — там, где сходятся границы соседних участков. По моим наблюдениям, такие сборища бывают очень кратковременными и не содержат элементов группового токования.
Гнездование.
Гнездо представляет собой ямку около 6 см глубиной, диаметром 20—22 см. Выстилка скудная, из небольшого числа сухих листьев и стеблей трав, среди которых может быть несколько перьев, оброненных насиживающей самкой. В первые дни после начала кладки выстилки вообще может не быть. Отмечены случаи, когда перед тем как снести первое яйцо самка обрамляет гнездовую ямку, еще не имеющую выстилки, несколькими веточками. Затем перед снесением каждого яйца, птица растаптывает на дне принесенные в клюве сухие листья, так что слой подстилки постепенно наращивается, а глубина гнездовой ямки уменьшается (Punnonen, 1954).
Для гнезда выбирается сухое место среди валежника, под нависшими ветвями елей или других деревьев, у самого ствола дерева в углублении между корнями, под густыми кустами, в выгнившей сердцевине старых низких пней, в зарослях папоротников. На севере ареала гнездо часто располагается в моховой подстилке. Оно обычно хорошо замаскировано и обнаружить его трудно. Известны случаи, когда гнездо устраивается в одном и том же месте два года подряд. Наконец, в очень редких случаях, как уже упоминалось, рябчик может гнездиться на деревьях, используя старые разрушенные гнезда врановых и хищных птиц. 4 таких случая известны для Финляндии (Punnonen, 1954). Под Ленинградом, в Тосненском районе, полная кладка из 10 яиц рябчика была найдена в старом гнезде врановой птицы (Дементьев, 1958). В таком же гнезде была найдена кладка рябчика на одном из островов р. Зеи (Заболотный, 1979). В последнем случае выводок рябчиков в конце концов благополучно оказался на земле и до поздней осени держался в районе гнезда, — таким образом, пуховые птенцы рябчика способны добираться до земли с высоты по крайней мере 5 м без особого вреда для себя.
Скорлупа яиц гладкая, желтовато-палевого цвета, с редкими бурыми пятнами различных размеров (от точек до пятен диаметром 5 мм), обычно резко очерченных. Количество их различно, они могут и вообще отсутствовать, а основная окраска яйца бывает то более темной, почти коричневой, то более светлой. Окраска яиц, как и у других тетеревиных, бывает особенно яркой в момент откладки, к концу же насиживания тускнеет и блекнет. Число яиц в кладке колеблется в пределах 3—14, обычно 7—9. Известно нахождение гнезд с кладками из 15, 18, 19 и даже 20 яиц (Мензбир, 1902; Punnonen, 1954). Вполне возможно, что в таких случаях мы имеем дело с гнездом, куда откладывали яйца две самки, тем более, что для этого вида известна совместная кладка даже с тетеревом (Киселев, 1978). Размеры яиц (103 штуки): 37—43x26—30 (Карелия; Ивантер, 1962); 37.9—42.4x26—29.8 мм; масса 13.1—18.8 г, в среднем 17.2 г (156 яиц, Вятско-Камское междуречье; Гайдар, 1974). Зависимость величины кладки от географического положения местности не прослеживается. Темп откладки яиц различен и, видимо, связан с возрастом самки, а также со степенью ее упитанности. Так, в одном случае самка отложила за 15 дней 9 яиц, другая за 13—14 дней — 10 яиц (Punnonen, 1954). По данным О. И. Семенова-Тян-Шанского, интервалы между откладкой яиц в одном из гнезд колебались от 26 до 39 ч, при этом время откладки постепенно сдвигалось от утра к вечеру.
В северной половине ареала кладка происходит в третьей декаде мая, в южной — во второй декаде. Значительно более ранние сроки на западе: в Белоруссии и в Прибалтике свежие полные кладки встречаются со второй половины апреля. Насиживание начинается с момента снесения последнего или предпоследнего яйца. Интенсивность насиживания варьирует индивидуально и в какой-то степени зависит от погоды. В первые дни самка может отлучаться на кормежки по 5 раз в сутки, но ближе к концу инкубации число кормежек сокращается до 2—3. Длительность отлучек на кормежку — 15—25 минут, хотя в отдельных случаях птица может отсутствовать до 35 мин. По различным данным, длительность насиживания составляет от 21 до 27 суток (Punnonen, 1954; Семенов-Тян-Шанский, 1959; Гайдар, 1974), что объясняется различной интенсивностью насиживания, особенно в первые дни.
Вылупление птенцов происходит в течение 8 ч (Долбик, 1961). Самка сразу же уводит птенцов от гнезда. Первое время выводок пасется на разреженных участках леса с невысоким травостоем или в зарослях папоротника, куда в достаточной степени проникают солнечные лучи. По некоторым данным (Нейфельд, 1958), птенцы уже на второй день жизни способны вспархивать на высоту до полутора метров. Несущую поверхность крыла у них в это время образуют первостепенные и второстепенные маховые перья, перекрывающие их основания большие кроющие крыла (каждое из кроющих несет на вершине две-три длинных пуховых бородки) и густой и длинный натальный пух, в свою очередь скрывающий основания кроющих перьев. Масса птенца в это время, составляет 12—14 г. По сравнению с птенцами глухаря рябчики в первые дни жизни требуют больше тепла. Даже в теплую погоду они через 5—6 мин пребывания на открытом воздухе начинают искать место для согревания, стремятся забраться под самку. В первые дни жизни птенцы склевывают корм только с растений, причем с нижней стороны стеблей и листьев, так, что клевки их направлены наискосок вверх (Данилов, 1975). Суточный прирост массы составляет в первые 10 дней 1.8—1.9 г, а первостепенные маховые перья вырастают на 2.6—3 мм. Рост этих перьев далее замедляется, составляя к концу развития пера 1.6 мм в сутки (Родионов, 1963б).
Появление первых выводков происходит на севере в середине—конце июня, в средней и южной частях ареала — в первой половине июня (в Монголии — в последней трети июня). В западных районах СССР выводки появляются с середины мая. Птенцы особенно прибавляют в массе в начале третьего месяца жизни — по 9 г в сутки и в конце этого месяца почти достигают массы взрослых птиц (Семенов-Тян-Шанский, 1959). На севере ареала это происходит в начале сентября, в средней, южной и западной частях — на 2—4 недели раньше. С поспеванием ягод выводки перекочевывают на ягодники, где и держатся большей частью до глубокой осени.
Самец в период насиживания держится обычно поблизости от гнезда (Punnonen, 1954; Гайдар, 1974), в насиживании не участвует. Выводок водит большей частью самка, но нередко вместе с самцом, который может заменять ее в случае гибели. Так, например, в заповеднике Денежкин Камень с выводком однажды был встречен только самец (Данилов, 1975). Самец может покидать выводок и в первые дни и значительно позже, но всегда раньше самки. Обычно он просто остается на своем участке, когда выводок с него откочевывает. Выводок начинает распадаться уже в августе, когда его начинают покидать молодые самцы.
Полный распад выводков, особенно поздних, может сильно затягиваться, вплоть до поздней осени, когда уже бывает трудно определить при встрече группы птиц — выводок это или образовавшаяся из распавшихся выводков стайка. Нередко несколько выводков, встречаясь на кормных местах, особенно на ягодниках, образуют скопления, насчитывающие до 30 птиц.
Осеннее токование, начинаясь с конца августа, длится в северных районах по октябрь, а в южных - вплоть до января (Хэнтэй; Козлова, 1930). Оно значительно менее оживленное, чем весеннее, и не сопровождается брачными играми и сперматогенезом.
Хозяйственное значение и охрана.
Будучи широко распространенным и обычным видом бореальных лесов Палеарктики, рябчик с незапамятных времен служил объектом промысла. Этому в немалой степени способствовало высокое качество его мяса. Хищнический промысел неоднократно приводил к полному истреблению рябчика в целых районах, но будучи оставленным в покое, рябчик неизменно восстанавливал свою численность в течение 2—3 лет. В конце XIX в. ежегодная добыча рябчиков только на рыночную продажу (в том числе и на экспорт) составляла ежегодно, по самым приблизительным подсчетам, около 3 млн. шт. (Мензбир, 1902). В настоящее время промысел этого вида еще ведется в ограниченных масштабах, но подавляющее большинство птиц добывается в процессе спортивной охоты или браконьерства и потребляется на месте. В последнее время численность вида заметно снизилась во многих местах, прежде всего на юге ареала в европейской части СССР из-за быстрого роста народонаселения и многократного усиления факторов беспокойства. Для охраны рябчика в таких условиях необходимо создание специальных заказников в наиболее удобных местах, что давало бы возможность птицам выводить птенцов без помех и поддерживать таким образом репродуктивный потенциал местных популяций. Само соседство с человеком рябчик переносит хорошо и нередко обитает буквально по окраинам деревень и вблизи от больших городов. В остальных же районах следует учитывать также важную роль, которую рябчик играет в питании таких ценных пушных видов, как соболь и куница.
Географическая изменчивость.
На всем протяжении обширного ареала изменчивость в окраске и размерах птиц сравнительно невелика и носит большей частью клинальный характер. Небольшим вариациям подвержены: тон окраски на верхней стороне тела, развитие белого цвета на брюхе, шейных, кроющих и плечевых перьях. Небольшим колебаниям подвержены также размеры птиц. Северные популяции отличаются более густым и длинным оперением плюсны, а также более густым оперением ноздрей и основания конька клюва. Особенно в этом отношении выделяются птицы с северо-востока ареала. Так, например, у отдельных особей с охотского побережья плюсна бывает оперена до самого основания.
Размеры и масса птиц на протяжении большей части ареала меняются сравнительно мало, будучи подвержены сильной индивидуальной изменчивости. Наиболее крупными размерами отличаются птицы из юго-восточных и особенно юго-западных популяций.
На всей северной и центральной частях ареала господствует серый тон окраски верха тела, с тонким поперечным полосатым рисунком на его передней части. По направлению к югу и западу основной тон верхней части тела становится более охристым, а поперечно-полосатый рисунок — более резким и грубым, но не везде. В сухих континентальных областях по югу ареала (Южный Урал, Хэнтэй) окраска птиц такая же, как и на севере, и даже еще более бледная. Наиболее интенсивным рыжим или даже красно-коричневым оттенком отличаются популяции на юго-западе ареала, особенно на Балканах (Bauer, 1960), и в меньшей степени — кое-где на юге (Алтай) и юго-востоке континентальной части ареала (юг Приморья, северная часть п-ова Корея). В наиболее континентальных частях ареала (Якутия, Колымский бассейн) у птиц в окраске довольно много белого цвета, особенно на брюхе, на плечевых и внутренних второстепенных маховых перьях.
В пределах ареала можно выделить следующие подвиды. Почти всю северную его часть и большую часть центральной населяет B. b. septentrionalis. Фенно-Скандинавию, Кольский п-ов и Прибалтику населяет номинативный подвид B. b. bonasia, по характеру окраски промежуточный между сибирским и западноевропейскими подвидами. В юго-западной части ареала по причине большей изоляции между отдельными популяциями изменчивость выражена более заметно. Здесь и выделена большая часть подвидов, но несмотря на сравнительно давнюю изоляцию отдельных популяций (порядка 5—10 тыс. лет) изменчивость не пошла дальше небольших изменений общего тона окраски и размеров. В настоящее время на территории Европы, на мой взгляд, можно считать реально существующими следующие подвиды: B. b. rupestris — верховья Одера, Эльбы Рейна, B. b. rhenana — Вогезы, Пфальцервальд, Хунсрюк, Эйфель, Таунас, Вестервальд и Зауерланд; крупный рыжий подвид B. b. styrlaca, синонимами которого, согласно ревизии Бауэра (Bauer, 1960), являются B. b. carpaticus и B. b. horicei, населяет леса Альп и Карпат. И, наконец, B. b. schiebeli с наиболее красно-рыжей окраской распространен в горах Балканского п-ова. Юг восточноевропейской части ареала занимает также довольно крупный, с хорошо выраженным рыжеватым оттенком верха B. b. volgensis. В восточной части ареала обитает наиболее светлый подвид B. b. kolymensis, причем между ним и сибирским подвидом имеется широкая переходная полоса. О-в Сахалин населяет B. b. yamashinai, который по чистоте серого цвета мантии приближается к предыдущему подвиду, но зоб и грудь имеют интенсивный охристый налет. Другой островной подвид, B. b. vicinitas, занимающий о-в Хоккайдо, также очень близок по окраске колымскому и сахалинскому подвидам, что указывает на вероятный путь проникновения рябчика на Хоккайдо через Сахалин. Юго-восток материкового ареала занимает подвид B. b. amurensis, который на границе с сибирским подвидом дает ряд переходов (некоторые из них были описаны как отдельные подвиды). Для этого подвида характерна интенсивная рыжая окраска зоба и охристый оттенок мантии.
Автор: Р. Л. Потапов. Источник: "Фауна СССР. Том III, вып. 1. Птицы. Отряд курообразные. Семейство тетеревиные.", 1985 г.
 Рябчик Северцова. Bonasa (Tetrastes) sewerzowi.
Рябчик Северцова. Bonasa (Tetrastes) sewerzowi. Троеперые рыбы Чёрного моря. Семейство Tripterygiidae.
Троеперые рыбы Чёрного моря. Семейство Tripterygiidae.Комментарии:
Нет комментариев :( Вы можете стать первым!
Добавить комментарий:
 Воротничковый рябчик. Bonasa umbellus.
Воротничковый рябчик. Bonasa umbellus. Рябчики.
Рябчики. Суточный бюджет энергии и времени тетеревиных птиц.
Суточный бюджет энергии и времени тетеревиных птиц. Использование снежного покрова тетеревиными птицами.
Использование снежного покрова тетеревиными птицами..") Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты).
Зимний сезон в жизни тетеревиных птиц (поведенческий и биоэнергетический аспекты).
